ÍNDICE
|
2. Pared celular 3. Matriz extracelular 12. Retículo endoplasmático 13. Aparato de Golgi 14. Lisosomas 15. Peroxisomas 16. Glioxisomas 17. Vacuolas 18. Mitocondrias 19. Plastos 1. Cloroplastos 20. Teorías sobre el origen de mitocondrias y cloroplastos 21. Ideas fundamentales 22. Prácticas 23. Tareas 24. Vídeos 25. Otras presentaciones 26. Cuestiones 27. Repaso 28. Actividades evaluables |
2. ESQUEMAS









3. PRESENTACIONES















Imágen célula procariota 2 3
Imágen célula eucariota animal 2 3
Imágen célula eucariota vegetal 2 3
Aparato de Golgi
Cloroplastos
Retículo endoplasmático
Lisosomas
Peroxisomas
Vacuolas
Cilios y flagelos
Citoesqueleto
Ribosomas
Cromosomas
Células en división
Formas celulares
5. CONTENIDOS ANIMADOS












6. EL ORIGEN DE LA VIDA:
Para explicar el origen de la vida, se suele aceptar la teoría de la sopa primitiva. Esta teoría describe como la vida se debió originar en los océanos, donde se dieron las condiciones adecuadas para que aparecieran moléculas sencillas en el agua y éstas se unieran formando compuestos más complejos en una especie de sopa o caldo. Estas moléculas entre las cuales estaban: proteínas, ácidos, azúcares, sales, grasas... se fueron más tarde uniendo en estructuras que fueron ensayos de lo que más tarde darían las células. Estos ensayos como esferas llenas de moléculas se llamaron coacervados. El autor de esta teoría fue Oparin pero muchos años más tarde ha sido comprobada en el laboratorio por otros científicos como Miller, Urey y Juan Oró.
|

ACTIVIDADES: 82 10 11 14 54



Durante el siglo XX, la teoría celular fue reafirmada y ampliada y es hoy uno de los conceptos unificadores más importantes de la biología. En su formulación actual, la teoría celular enuncia:
1) Los seres vivos están formados por células y productos celulares.
2) Las células se originan a partir de otras células.
3) Las reacciones químicas del organismo vivo tienen lugar dentro de células.
4) Las células contienen la información hereditaria de los organismos que integran y esta información se transmite de la célula madre a la célula hija.
Todas aquellas características que se hacen evidentes en un organismo complejo y nos permiten reconocerlo como un ser vivo, están presentes en cada una de las células que lo componen.
Las características de las células son:
Que la superficie celular, a través de la cual se realizan los intercambios con el medio, dé abasto para suministrarle a la célula los nutrientes necesarios y permitirle la eliminación de sus desechos. Cuanto mayor es el volumen de un cuerpo, proporcionalmente menor resulta su superficie. Tómese este simple ejemplo: un cubo A de 30 micrómetro de lado tiene una superficie de 900 micrómetros cuadrados y un volumen de 2700 micrómetro cúbico. Un cubo B de 10 micrómetros de lado tiene una superficie de 900 micrómetros cuadrados cada uno de los 27 cubos pequeños, lo que determina una superficie total de 27000 micrómetros cuadrados y un volumen de 1000 micrómetros cúbicos cada cubo pequeño para un total de 2700 micrómetros cúbicos. Si la superficie de estos cuerpos tuviera que ser utilizada para realizar intercambios, como ocurre con las células, el cuerpo B sería más eficiente que A, ya que dispone de una superficie relativamente mayor para su volumen.

Que el volumen celular sea lo suficientemente pequeño para que las moléculas que participan del metabolismo puedan llegar de una parte a otra de la célula en un tiempo breve.
Unidades de longitud utilizadas en Biología celular


El tamaño de las células es muy variable, la mayoría miden entre 5 y 50 micras. Una micra (μ) o micrómetro (μm) equivale a 0,000001 m. Algunas células son visibles a simple vista como los óvulos de las gallinas (huevos).
Un microscopio óptico es un
microscopio basado en lentes ópticos. También se le conoce como microscopio de luz, (que utiliza luz o "fotones") o microscopio de campo claro. El desarrollo de este aparato suele asociarse con los trabajos de Anton van Leeuwenhoek. Los microscopios de Leeuwenhoek constaban de una única lente pequeña y convexa, montada sobre una plancha, con un mecanismo para sujetar el material que se iba a examinar (la muestra o espécimen). Este uso de una única lente convexa se conoce como microscopio simple, en el que se incluye la lupa, entre otros aparatos ópticos.


Un microscopio electrónico es aquél que utiliza electrones en lugar de fotones o luz visible para formar imágenes de objetos diminutos. Los microscopios electrónicos permiten alcanzar ampliaciones hasta 5100 veces más potentes que los mejores microscopios ópticos, debido a que la longitud de onda de los electrones es mucho menor que la de los fotones "visibles".
Un microscopio electrónico, como el de la imagen, funciona con un haz de electrones generados por un cañón electrónico, acelerados por un alto voltaje y focalizados por medio de lentes magnéticas (todo ello al alto vacío ya que los electrones son absorbidos por el aire). Un rayo de electrones atraviesa la muestra (debidamente deshidratada y en algunos casos recubierta de una fina capa metálica para resaltar su textura) y la amplificación se produce por un conjunto de lentes magnéticas que forman una imagen sobre una placa fotográfica o sobre una pantalla sensible al impacto de los electrones que transfiere la imagen formada a la pantalla de un ordenador. Los microscopios electrónicos producen imágenes sin ninguna clase de información de color, puesto que este es una propiedad de la luz y no hay una forma posible de reproducir este fenómeno mediante los electrones; sin embargo, es posible colorear las imágenes posteriormente, aplicando técnicas de retoque digital a través del ordenador.



ANIMACIONES













ANIMACIONES


9.1. Células procariotas. Son las células que no tienen núcleo, es decir son las que presentan su ADN más o menos condensado en una región del citoplasma pero sin estar rodeado de una membrana. El ejemplo más importante de células procariotas son las bacterias. Son células muy sencillas, sus orgánulos prácticamente sólo son los ribosomas, los mesosomes (unos orgánulos exclusivos de estas células) y algunas también tienen unos flagelos muy sencillos.










ACTIVIDADES: 10 1 2 3 5 6 8 8 35 36
9.2. Células eucariotas. Son las células que tienen núcleo, es decir son las que presentan su ADN rodeado de una membrana. Tienen estructura eucariota las células de los animales, plantas,algas, hongos y protozoos.
La célula eucariota. Es puede definir como una estructura biológica constituida por tres partes denominadas membrana plasmática, citoplasma y núcleo , y que es capaz de realizar las tres funciones vitales. La célula eucariota es la unidad estructural y funcional de todos los organismos pluricelulares. Presenta formas y tamaños muy diferentes. Generalmente tienen una medida de unos unos 0,020 mm, pero algunas células eucariotas, como la yema del huevo de gallina, tienen más de un centímetro de diámetro

9.2.3. DIFERENCIAS ENTRE CÉLULAS ANIMALES Y VEGETALES
10. ORGÁNULOS CELULARES Y SU FUNCIÓN
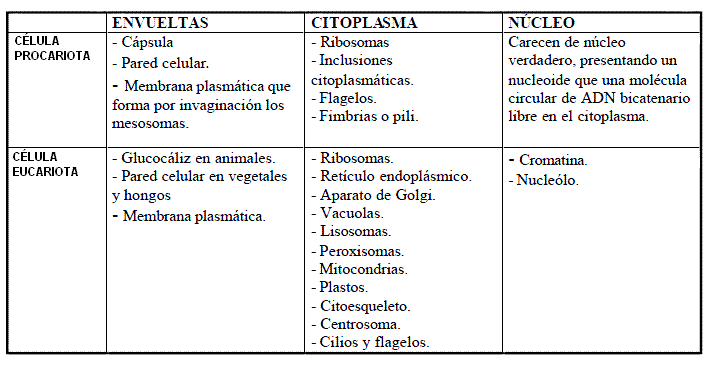 11. LAS ENVOLTURAS CELULARES
11. LAS ENVOLTURAS CELULARES
Las envolturas celulares son capas que separan el medio interno del exterior. Son: la membrana plasmática, que poseen todas las células (eucariotas y procariotas) y las membranas de secreción (que pueden faltar). Son membranas de secreción: la matriz extracelular (de animales), la pared vegetal (de vegetales) y la pared bacteriana (en bacterias).
11.1. La membrana plasmática
La membrana plasmática constituye el límite entre el citoplasma y el medio en el que se encuentra la célula y entre los orgánulos celulares y el citosol (hialoplasma), de manera que las biomembranas dividen al interior de la célula en numerosos compartimentos. De esta manera se explica que las membranas puedan constituir el 60 % del citoplasma celular.
el 60 % del citoplasma celular.
Posee un espesor de 75 Ǻ (ángstrom). Al microscopio electrónico se presenta como una triple capa. Dos bandas oscuras externas de 20 Ǻ separadas por una interna de color claro de 35 Ǻ. En ocasiones, las láminas externas más oscuras pueden tener espesores diferentes.
Cualquier tipo de célula presenta este tipo de membrana de tres capas. Debido a su carácter generalizado se le denomina unidad de membrana (o membrana unidad). También se encuentra formando la envoltura exterior de muchos orgánulos como los cloroplastos, mitocondrias o vacuolas.



11.1.1. Composición química
Las membranas biológicas son conjuntos laminares constituidos aproximadamente por un 40 % de lípidos y un 60 % de proteínas principalmente; asociados a los lípidos y las proteínas también se encuentran oligosacáridos.
1. Lípidos. Son esencialmente anfipáticos, o sea que sus moléculas poseen un polo hidrófilo y un polo hidrófobo. Los más abundantes (en los glóbulos rojo humano) son los fosfolípidos (55 % del total de los lípidos), el colesterol (25%) y otros lípidos, glucolípidos y ácidos grasos(20 %) que son enteramente hidrófobos.
Los lípidos, cuando se encuentran en un medio acuoso, se disponen formando una doble capa, la bicapa lipídica, con las zonas polares (hidrófilas) orientadas hacia el exterior, y las zonas apolares (hidrófobas) protegiéndose mutuamente.
 La bicapa lipídica no es una estructura rígida, sino que sus componentes se mueven en ella con libertad (Fig. 2), confiriéndole fluidez. Las moléculas de lípidos, más móviles, pueden girar sobre sí mismas (rotación) o intercambiar su posición con otras moléculas de la misma monocapa (difusión lateral). Es poco frecuente el intercambio entre moléculas situadas en monocapas distintas (flip-flop).
La bicapa lipídica no es una estructura rígida, sino que sus componentes se mueven en ella con libertad (Fig. 2), confiriéndole fluidez. Las moléculas de lípidos, más móviles, pueden girar sobre sí mismas (rotación) o intercambiar su posición con otras moléculas de la misma monocapa (difusión lateral). Es poco frecuente el intercambio entre moléculas situadas en monocapas distintas (flip-flop).
▪ El colesterol contribuye grandemente a la fluidez de la bicapa, debido a que con su pequeño tamaño dificulta el establecimiento de interacciones hidrofóbicas entre las colas apolares de los lípidos de membrana.



2. Las proteínas se disponen intercaladas o adosadas a la bicapa de lípidos, y son de diverso tamaño y naturaleza.
Por su afinidad a los lípidos de la membrana hay dos tipos:


3. Oligosacáridos. Se asocian a lípidos (glucolípidos) o a proteínas (glucoproteínas).


La situación de las proteínas (integrales o periféricas) en la membrana la determina su afinidad por el agua o los lípidos. Los azúcares (oligosacáridos), asociados a proteínas (glucopropteínas) o lípidos (glucolípidos), se sitúan en la cara extracelular de la membrana. A esta cubierta se la llama glucocáliz o cubierta celular.
Según lo expuesto, el modelo de mosaico fluido de membrana sostiene:
▪ Los lípidos y proteínas que forman la membrana plasmática constituyen un mosaico molecular. Están dispuestas unas junto a otras como las piezas de un mosaico.
▪ Los lípidos y proteínas pueden desplazarse en el plano de la bicapa lipídica: las membranas son fluidas.
▪ Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares. La asimetría se debe a que la presencia de oligosacáridos está restringida a la superficie de la cara externa y, además, la distribución de los lípidos en una y otra monocapa no es simétrica.
11.1.3. Funciones de la membrana plasmática
● Frontera física entre dos medios. Esto permite no sólo la separación del interior de la célula con respecto al medio exterior (intra y extracelular), sino también la formación de compartimentos en el interior de la célula eucariótica.
● Facilita que ocurran, de manera simultánea, pero sin mezclarse, una gran diversidad de reacciones químicas en sus diferentes orgánulos.
● La bicapa lipídica es una eficaz barrera para evitar el paso de sustancias hidrófilas (se evita la pérdida de sustancias intracelulares).
● Asegura el intercambio y transferencia de sustancias e información con el exterior y con otras células. La naturaleza lipídica de la membrana determina el tipo de sustancias que pueden atravesarla y, además, las proteínas que la forman, pueden intervenir de manera activa facilitando o impidiendo el transporte de esas sustancias.
Otras funciones de las diferentes membranas, determinadas por la presencia de proteínas específicas, son:
● Factores de reconocimiento celular. Conforman la “identidad antigénica” de cada individuo debido a que las proteínas específicas de la membrana celular constituyen una combinación única en cada individuo, que permite ser reconocida por las defensas inmunitarias.
● Receptores hormonales y de otras informaciones. Control del flujo de información entre las células y el medio, reciben la información que llega del medio, gracia a la existencia de a) receptores específicos de neurotransmisores y de hormonas, y b) el potencial de membrana responsable de la sensibilidad celular.
● Desempeñar funciones especiales gracias a las diferenciaciones que presentan algunas: invaginaciones (aumento de la superficie de intercambio), desmosomas (zonas de unión con otras células), etc.
11.1.3.1. Transporte a través de la membrana plasmática.
Las membranas son barreras de permeabilidad muy selectiva. Los mecanismos que utilizan las células para permitir el paso de sustancias varían en función de que se trate de moléculas pequeñas, que puedan atravesarla, o de moléculas más grandes, que deban ser englobadas y posteriormente liberadas por la propia membrana:
1. Moléculas pequeñas. La permeabilidad de las membranas celulares es altamente selectiva. El paso a su través de moléculas e iones está controlado por mecanismos de transporte específicos. Existen dos mecanismos básicos de transporte: el activo y el pasivo.

Un ejemplo es la bomba de sodio-potasio, que mantiene el potencial electroquímico a ambos lados de la membrana de las células animales (bombea Na+ hacia el exterior de la célula y K+ hacia el interior).


III.- Uniones intercelulares: para mantener adheridas y comunicadas células vecinas.
• Uniones occludens (impermeables). No dejan espacio entre las células e impiden el paso de sustancias, actuando a modo de barrera. Son frecuentes entre las células epiteliales.
• Uniones comunicantes. En ellas existe un pequeño espacio intercelular, con lo que las membranas no llegan a contactar y permite el paso de pequeñas moléculas entre dos células vecinas. Se pueden distinguir dos tipos:
Sinapsis, se realizan entre dos neuronas separadas por la hendidura sináptica.
Uniones en hendidura o de tipo gap (del ingles, hendidura), dejan entre sí una hendidura lo suficientemente ancha como para permitir el paso entre ellas de moléculas relativamente grandes.

• Uniones adherentes (desmosomas). El espacio intercelular aumenta y en la cara interna de la membrana plasmática se sitúa un material denso, denominado “placa”, hacia el que se dirigen haces de filamentos. Estas uniones se localizan en aquellos tejidos que se encuentran sometidos a esfuerzos mecánicos.






11.2. PARED CELULAR VEGETAL.
La pared celular es una forma especializada de matriz extracelular (segregada por la célula y excreta al exterior de la membrana plasmática), que se encuentra adosada a la membrana plasmática de las células vegetales, y que se caracteriza por su alto contenido encelulosa, lo que la hace ser gruesa, rígida y organizada.
11.2.1. Composición química de la pared vegetal
Como ya hemos señalado, esta formada principalmente por celulosa (homopolisacárido que se origina por la unión β (1→4) de la D-glucosa), pero también por: Hemicelulosa, Pectinas, Sales minerales y Agua
11.2.2. Estructura de la pared vegetal
Está constituida por tres capas, cada una con distinta composición y características. Desde fuera hacia dentro son:
 I.- Lámina media: es la capa más externa y es común a las dos células adyacentes. Es delgada y flexible, y está compuesta principalmente por Pectinatos de calcio. Se encarga de mantener unidas las distintas células en los tejidos vegetales.
I.- Lámina media: es la capa más externa y es común a las dos células adyacentes. Es delgada y flexible, y está compuesta principalmente por Pectinatos de calcio. Se encarga de mantener unidas las distintas células en los tejidos vegetales.
II.- Pared primaria: capa relativamente delgada y semirrígida, típica de las células jóvenes, recién divididas (plantas en crecimiento). Está formada por celulosacon una abundante matriz hemicelulósica.
III.- Pared secundaria: capa muy gruesa formada por varias subcapas de celulosa, en cada una de las cuales las fibras de celulosa se disponen con distinta orientación, lo cual le da a la pared una gran rigidez y resistencia. La pared secundaria sólo se presenta en células maduras o ya muertas. Precisamente el grosor de la capa de celulosa hace que el citoplasma se vaya "asfixiando", y la célula acabe por morir.



11.2.3. Funciones de la pared vegetal
▪ Constituyen un exoesqueleto que protege a la célula, le da forma y le confiere resistencia, pero sin impedir su crecimiento.
▪ Es la responsable de que la planta se mantenga erguida.
▪ Impide que la célula se rompa, ya que interviene activamente en el mantenimiento de la presión osmótica intracelular.
▪ Permite la comunicación entre células adyacentes y con el exterior, para el intercambio de nutrientes y de información. Existen unos orificios que atraviesan la pared llamados punteaduras que se sitúan al mismo nivel en células vecinas. Estas punteaduras son atravesadas por puentes citoplasmáticos o plasmodesmos, que son prolongaciones del retículo endoplasmático.
11.3. MATRIZ EXTRACELULAR
Se encuentra por fuera de la membrana celular y desempeña un papel muy importante en la vida de la célula. Esta constituida por compuestos que segregan localmente la propia células.
El glucocálix (conjunto de cadenas de oligosacáridos) aparece en la cara externa de la membrana celular de muchas células animales. Tiene funciones de reconocimiento celular indispensables para la fecundación, reconocimiento de la célula a parasitar de virus y bacterias, adhesión de células para formación de tejidos y recepción de antígenos específicos para cada célula.
Su estructura consiste en una fina red de fibras de proteína inmersa en una estructura gelatinosa de glucoproteínas hidratadas, la sustancia fundamental amorfa.
En su composición química hay fundamentalmente: colágeno, elastina, fibronactina, glucoproteínas.
La función es primordialmente servir de unión y nexo en los tejidos conectivos, cartilaginoso y conjuntivo. Puede acumular sales, originando tejido óseo o quitina y dando lugar a exoesqueletos.



ANIMACIONES


CUESTIONES: 1 2 3 4 5 6 4
12. RETÍCULO ENDOPLASMÁTICO.
Está formado por un conjunto de membranas que limitan cavidades cerradas o cisternas de distintas formas: sáculos aplanados,vesículas globulares o tubos de aspecto sinuoso que delimitan un espacio interno, lumen o espacio cisternal contenido en el interior del retículo endoplásmatico y el espacio citosólico en el exterior del retículo. Estas cavidades se comunican a menudo entre ellas y forman una red (retículo = pequeña red) característica de las células eucariotas.
retículo. Estas cavidades se comunican a menudo entre ellas y forman una red (retículo = pequeña red) característica de las células eucariotas.
Sacos del retículo, por lo tanto, dobles membranas, separan el material nuclear del resto del citoplasma. Esta doble membrana está atravesada por una serie de poros que permiten el paso de grandes moléculas (ARN, subunidades ribosómicas, etc.).
Podemos distinguir dos tipos de retículo:
1.1. Retículo endoplasmático rugoso, que posee ribosomas adheridos a la cara de la membrana que da al hialoplasma (espacio citosólico). Se encuentra muy desarrollado en aquellas células que participan activamente en la síntesis de proteínas.
Dentro de sus funciones destacan: la síntesis de proteínas por parte de los ribosomas que pueden ser almacenadas en el del retículo donde maduran con frecuencia sufren una glucosilación de las proteínas (se unen a un glucido, glucoproteínas) y son transportadashacia otros orgánulos (aparato de Golgi, lisosomas), a la membrana celular o las propias membranas del retículo.
1.2. Retículo endoplasmático liso, constituido por finos túmulos o canalículos interconectados y cuyas membranas se continúan con las del REr, pero sin llevar adheridos ribosomas.
 Sus funciones son: la síntesis lípidos de membrana (fosfolípidos, colesterol, etc.). Asimismo, en el retículo tiene lugar la detoxificación de aquellas sustancias perjudicialespara la célula producidas por su actividad vital o procedente del exterior (insecticidas, herbicidas, medicamentos, etc.). Las toxinas que proceden del medio externo llegan a nuestras células, son transformadas por este orgánulo en moléculas cuya toxicidad se ha visto reducida y que son de esta manera más fácilmente eliminadas.
Sus funciones son: la síntesis lípidos de membrana (fosfolípidos, colesterol, etc.). Asimismo, en el retículo tiene lugar la detoxificación de aquellas sustancias perjudicialespara la célula producidas por su actividad vital o procedente del exterior (insecticidas, herbicidas, medicamentos, etc.). Las toxinas que proceden del medio externo llegan a nuestras células, son transformadas por este orgánulo en moléculas cuya toxicidad se ha visto reducida y que son de esta manera más fácilmente eliminadas.
Gracia a la acción conjunta de ambos retículos queda completa la síntesis de los componentes básicos de toda membrana celular: proteínas y lípidos.




13. APARATO DE GOLGI.
El aparato de Golgi es un orgánulo membranoso que forma un sistema de cavidades aplanadas dispuestas ordenadamente. Cada una de estas cavidades recibe el nombre de sáculo. El conjunto de sáculos apilados recibe el nombre de dictiosoma. Además, pueden observarse toda una serie de vesículas más o menos esféricas a ambos lados y entre los sáculos relacionadas con el transporte de proteínas y lípidos desde y hacia el A. de Golgi.
El conjunto de todos los dictiosomas y vesículas celulares constituye el aparato o complejo de Golgi. Se encuentra más desarrollado cuanto mayor es la actividad celular y en las células secretoras.
El aparato de Golgi tiene dos caras distintas, la cara “cis”, “externa” o de formación y la cara “trans”, “interna” o de maduración.
Los dictiosomas no son estructuras celulares fijas, sino que se están renovando constantemente. Se forman a partir del retículo endoplásmatico. Este emite unas prolongaciones que se separan de él formando vesículas de transición (estas encierran en su interior el contenido de las cavidades del retículo, fundamentalmente proteínas, que se han fabricado en los ribosomas del retículo endoplasmático rugoso y cuyo destino es la secreción). Al unirse posteriormente dichas vesículas entre sí dan lugar a un sáculo (cara cis o de formación). Por otra parte, las cavidades golgianas opuestas (cara trans o de maduración) se fragmentan para formar vesículas cargadas de productos de secreción.
Funciones. El aparato de Golgi funciona como una planta “empaquetadora” y “distribuidora” de los productos fabricados por el R. E.
● Embalaje y secreción. Las sustancias fabricadas en el R. E. para ser secretadas se incorporan a la cara “cis” o de formación del aparato de Golgi y se desplazan progresivamente hacia la cara “trans” o de maduración, desde donde se incorporan a las vesículas de secreción. Las vesículas se adosan a la membrana y por un proceso de exocitosis vierten su contenido al medio extracelular.
● Este proceso genera un reciclaje permanente de la membranacompensado por los mecanismos de endocitosis. Entre los productos secretados tienen especial interés los que forman la matriz extracelular que, en los vegetales, origina la pared celular.
● No todos los productos empaquetados por el aparato de Golgi tienen como destino el exterior de la célula. Por ejemplo, algunas vesículas, que contienen abundantes enzimas hidrolíticos, se transforman en lisosomas y permanecen en el citoplasma celular.
● En los retículos endoplasmáticos se sintetizan los componentes moleculares de las membranas (lípidos en el liso y proteínas en el rugoso) y el contenido de sus cavidades. Del retículo se desprenden vesículas que se incorporan a los sacos del aparato de Golgi, donde sus componentes moleculares experimentan modificaciones, glucolisaciones, por la adición de oligosacáridos, que se unen a proteínas (glucolisación de proteínas: glucoproteínas de membrana, anticuerpos) y a lípidos (glucolisación de lípidos: glucolípidos de membrana).




ANIMACIONES





14. LISOSOMAS.
Los lisosomas son orgánulos celulares, globulares que presentan formas muy diversas. Se originan a partir del retículo endoplásmico o del aparato de Golgi, de la misma manera que se originan las vesículas de secreción.
Constituyen el aparato digestivo de la célula. Contienen enzimas hidrolíticas (fosfatasa ácida, proteasa,...) que catalizan la hidrólisis o digestión de las macromoléculas; la membrana que limita a cada lisosoma impide que la célula sea digerida por estos enzimas líticos.
 Se encuentran en las células eucariotas tanto animales como vegetales, la estructura de estos orgánulos es muy distinta de un tipo celular a otro e incluso en una misma célula. Este polimorfismo hace que no sea posible caracterizar a los lisosomas bajo criterios estrictamente estructurales y tan sólo la demostración de actividades líticas permite poner de manifiesto la naturaleza lisosomica de una inclusión celular.
Se encuentran en las células eucariotas tanto animales como vegetales, la estructura de estos orgánulos es muy distinta de un tipo celular a otro e incluso en una misma célula. Este polimorfismo hace que no sea posible caracterizar a los lisosomas bajo criterios estrictamente estructurales y tan sólo la demostración de actividades líticas permite poner de manifiesto la naturaleza lisosomica de una inclusión celular.
A pesar de esta diversidad anatómica, los lisosomas se clasifican en dos categorías:
Lisosomas primarios, recién formados a partir del Ap. de Golgi o del R.E., de forma ovalada o redondeada, que no han intervenido en ningún proceso de digestión y sólo contienen enzimas hidrolíticos.
Lisosomas secundarios, de forma variable (depende de la naturaleza de los sustratos y del estado de digestión en el que se encuentren), están implicados en algún proceso de digestión. Se forman a partir de la unión de los lisosomas primarios con endosomas que contienen sustratos procedentes del medio externo (vacuolas heterofágicas o digestivas) o interno (vacuolas autofágicas).
Funciones.
A.- Digestión celular
■ Vacuolas heterofágicas o digestivas (heterofagia) con sustratos procedentes del medio externo, cuya finalidad es alimenticia y defensiva.
Las partículas procedentes del medio externo penetran en la célula, mediante un proceso de invaginación de la membrana celular, que se separa de ésta, formándose una vacuola alimenticia o vacuola de endocitosis. Los lisosomas primarios se unen a esta vacuola, dando lugar a un orgánulo único, la vacuola digestiva o lisosoma secundario, que contiene no sólo enzimas hidrolíticos, sino también moléculas que va a digerir, por lo que se considera a esta vacuola el estómago celular.

Estas moléculas, una vez que son digeridas, pasan al citoplasma celular. Sin embargo, no todas las sustancias son digeridas y por tanto no pueden ser aprovechadas por la célula. A estas partículas no digeridas se las llama productos residuales, siendo expulsados al exterior de la célula después de que la vacuola digestiva se ha transformado en vacuola fecal.
■ Vacuolas autofagicas (autofagia) si el sustrato lo constituyen componentes de la propia célula. Este mecanismo interviene en los procesos de desarrolla y asegura la nutrición en condiciones desfavorables.
Los lisosomas producen la hidrólisis de moléculas u orgánulos de la propia célula, tal es el envejecimiento celular, estados de ayuno en los que la digestión de sus constituyentes celulares propician los alimentos que la células necesitan. Cuando esto ocurre, el proceso recibe el nombre de autofagia, que significa comerse a sí mismo.
B. Digestón extracelular. En algunas ocasiones, los lisosomas vierten su contenido al exterior de la célula, donde tiene lugar la digestión (digestión extracelular).


15. PEROXISOMAS.
Son orgánulos presentes en casi todas las células eucariotas. Al igual que los lisosomas, son compartimentos vesiculares que contienen enzimas, pero éstas, a diferencia de las de los lisisomas, intervienen en reacciones oxidativas en las que se consume gran cantidad de oxígeno.
En esta reacción, RH2 representa una molécula orgánica que se oxida. Como resultado de este tipo de oxidaciones se forma como producto H2O2, peróxido de hidrógeno (agua oxigenada) , auténtico veneno para la célula; sin embargo, éste no afecta a la célula, ya que los peroxisomas contienen una elevada cantidad de la enzima catalasa que cataliza la reacción:
De esta manera desaparece el peróxido de hidrógeno en el mismo orgánulo en el que se ha producido, no afectando
al citosol que le rodea. Estos orgánulos se forman por gemación de la membrana del retículo endoplasmático y suelen localizarse junto a él.
16. GLIOXISOMAS.
Orgánulos exclusivos de las células vegetales. Son parecidos a los peroxisomas, y se encargan, durante la germinación, de transformar los lípidos almacenados en la semilla en glúcidos necesarios para el crecimiento del embrión hasta que la plántula pueda realizar la fotosíntesis. Poseen enzimas del ciclo del ácido glioxílico que es una variante del ciclo de Krebs de las mitocondrias que permite sintetizar azúcares a partir de grasas. Es indispensable en semillas en germinación.
SISTEMA DE ENDOMEMBRANAS
Es el conjunto de membranas citoplasmáticas internas: fundamentalmente RE, dictiosomas, membrana plasmática y vacuolas, y constituye una unidad funcional.
Son estructuras móviles, continuamente cambiantes, con el RE como fuente de crecimiento de las mismas. No participan en los plástidos ni las mitocondrias.




17. VACUOLAS
Almacenamiento de sustancias de reserva. Vesículas de forma globular y de tamaño variable, destinadas a almacenar todo tipo de sustancias.
Se forman a partir de muchos tipos de orgánulos membranosos:
La estructura de las vacuolas es muy sencilla: consiste solamente en un territorio celular rodeado de una membrana. Al conjunto de vacuolas de una célula se denomina Vacuoma.
Las células vegetales presentan vacuolas de gran tamaño, que a veces desplazan a la periferia todo el citoplasma celular y el núcleo, y son escasas en número. Esta vacuola recibe el nombre de vacuola central. En cambio, las células animales poseen numerosas vacuolas de pequeño tamaño, habitualmente denominadas vesículas.
Funciones.
• Almacenar sustancias de reserva:
• Servir de vehículo de transporte de sustancias entre orgánulos citoplasmáticos (vesículas de transición, de secreción, etc.) y entre el exterior e interior de la célula (vacuolas o vesículas de endocitosis).
• Regulan la presión osmótica de la célula. Las vacuolas pueden llenarse o vaciarse de agua para mantener constante la concentración del citoplasma.
• Vacuolas pulsátiles, es un caso especial son las que eliminan el exceso de agua citoplasmática en células de organismos dulceacuícolas (que son hipertónicos respecto al medio).


CUESTIONES: 1 2

18. MITOCONDRIAS.
Son orgánulos presentes en el citoplasma de todas las células eucarióticas (aeróbicas). Su forma se asemeja a un cilindro alargado, difíciles de observar al microscopio óptico, al que aparecen como palitos o bastoncitos alargados. Lo más frecuente es que las mitocondrias estén dispersas en el hialoplasma, y su número depende del tamaño de la célula: unas pocas en las levaduras y de 1000 a2000 en una célula hepática. Son las encargadas de la obtención de la energía mediante la respiración celular, proceso de oxidación en el que intervienen las ATP sintetasas. La energía obtenida se guarda en forma de ATP.
Ultraestructura.
La observación al microscopio electrónico pone de manifiesto que cada mitocondria está separada del hialoplasma por una membrana continua de 60 Å (anstrom) de espesor, que es la membrana mitocondrial externa. Esta membrana externa está rodeada interiormente por una segunda membrana, igualmente continua y de 60 Å de espesor, que es la membrana mitocondrial interna. La membrana interna forma repliegues orientados hacia el interior de la mitocondria y son las llamadas crestas mitocondriales (por lo que su superficie es 5 veces mayor que la de la membrana externa). Las dos membranas mitocondriales delimitan dos compartimentos diferentes: un primer compartimento situado entre la membrana mitocondrial externa y la interna que es el espacio intermembrana; un segundo compartimento limitado por la membrana mitocondrial interna que es la matriz mitocondrial. En la matriz encontramos ADN y ribosomas mitocondriales.


1º. Oxidaciones respiratorias: Cuya finalidad es obtener energía para la célula a partir de materia orgánica. Consisten en la degradación completa de moléculas orgánicas a CO2 para liberar energía y formar ATP. Tienen lugar en dos series de procesos, el ciclo de Krebs, la beta oxidación de los ácidos grasos y la cadena de transporte de electrones, que suceden, respectivamente, en la matriz y en la membrana mitocondrial interna.
2º. Producción de metabolitos precursoras para la biosíntesis de macromoléculas, se sintetizan en el hialoplasma.
3º. Síntesis de proteínas mitocondriales, ya que, poseen la maquinaria y la información necesaria para ello.

19. PLASTOS.
Los Cloroplastos están englobados en los Plastos o Plastidios, conjunto de orgánulos exclusivos de células vegetales que tienen un origen común. Poseen, al igual que las mitocondrias, información genética propia, ribosomas y una doble membrana. Hay tres tipos de plastos, aunque su estructura es muy semejante:
● Leucoplastos: (leuco = blanco). En partes no verdes del vegetal - las subterráneas, por ejemplo- entre ellos destacan losamiloplastos que acumulan almidón en los tejidos de reservas.
● Cromoplastos: (cromo = color). Dan color rojo, anaranjado o amarillo a muchos frutos, flores y otros órganos. Acumulan pigmentos fotosintéticos secundarios (xantofilas y carotenos) y reservas nutritivas.
● Cloroplastos: (cloro = verde). Son de color verde debido a la Clorofila que contienen. Se encargan de realizar la Fotosíntesis, captando energía luminosa.
19.1. Cloroplastos
Los cloroplastos se localizan en células vegetales fotosintéticas y, en los vegetales superiores, tienen forma lenticular (de lenteja). Su número suele ser de unos 40 por célula. Son de color verde debido a su elevado contenido en clorofila.
 Funciones de los cloroplastos.
Funciones de los cloroplastos.
Una de ellas es la síntesis de proteínas cloroplásticas, ya que, como hemos visto, poseen la maquinaria y la información necesaria para ello.
Pero la su principal función es llevar a cabo la Fotosíntesis, que consiste en la formación de materia orgánica a partir de compuestos inorgánicos (H2O, CO2, NO3=, SO4=, PO43-...), que reducen gracias a la energía captada por los pigmentos fotosintéticos.
En los tilacoides se encuentran los pigmentos captadores de luz, y la cadena transportadora de electrones (o cadena fotosintética), donde el ADP sé fosforila y pasa a ATP. Al ser la luz, digamos, el motor de este proceso, se le da el nombre de fotofosforilación. En los tilacoides se produce también la reducción del NADP+, que pasa a NADPH + H+. Ambos procesos constituyen lo que se conoce como fase lumínica de la fotosíntesis.
La energía acumulada, tanto en el ATP como en el NADPH + H+, será utilizada en la fase oscura para la fijación del CO2 a moléculas orgánicas. Esta fase oscura de la fotosíntesis tiene lugar en el estroma del cloroplasto.
Como ocurre en la mitocondria, en los cloroplastos se encuentran también esferas de ATP-sintetasa (o ATPasa), situadas en lamembrana interna del cloroplasto que mira hacia el estroma y en la membrana de los tilacoides. La ATPasa cataliza la reacción de formación de ATP
.

20. TEORÍAS SOBRE EL ORIGEN DE MITOCONDRIAS Y CLOROPLASTOS.
Recordemos que los organismos vivos más antiguos sobre la Tierra eran células procarióticas (3.500 millones de años), habiendo aparecido mucho más tarde las células eucarióticas (700 m.a. tienen los restos más antiguos encontrados de estos organismos). Por tanto, lo único que se puede afirmar es que probablemente, las células eucariotas evolucionaron a partir de procariotas entre los 3.500 y los700 m.a.
Actualmente existen dos teorías que tratan de explicar el posible origen de la célula eucariota.
La Teoría endógena propone que la célula eucariótica es el resultado del aumento de tamaño de la procariótica, junto a una progresiva diferenciación interna, cuyo resultado ha sido el alto grado de complejidad que posee.
La Teoría endosimbiótica, propuesta por la bióloga Lynn Margulis, considera que el alto grado de complejidad se debe a asociaciones entre células que en principio eran independientes entre sí. Este tipo de asociación debió consistir en una simbiosis, relación en la que ambas especies asociadas resultan beneficiadas.
En esta asociación las premitocondrias, que serían bacterias aerobias, conseguirían la oxidación de los alimentos, y las células hospedadoras obtendrían una ganancia en energía. A su vez, los precloroplastos, antiguas cianobacterias, suministrarían alimentos a las células hospedadoras mediante fotosíntesis, y obtendrían de ellas moléculas simples necesarias para realizar dicha función. La asociación de procariotas del tipo de espiroquetas daría lugar, por otro lado, a centríolos, cilios y flagelos.
Después de millones de años de dependencia mutua con las células hospedadoras, estos procariotas simbiontes perderían su independencia, conservando únicamente algunos vestigios de su forma de vida anterior.
De acuerdo con esta hipótesis, las antiguas bacterias habrían entrado al interior de la célula hospedadora por endocitosis, encerradas en unas vesículas endocíticas; esta circunstancia explicaría la doble membrana que poseen tanto mitocondrias como cloroplastos.
Esta teoría endosimbiótica se ve avalada por los siguientes hechos:
1. Tanto mitocondrias como cloroplastos contienen ADN del tipo de células procariotas, no asociado a proteínas histónicas.
2. Los dos orgánulos poseen ribosomas con los que sintetizan sus propias proteínas, también de tipo procariótico (70 S).
3. Ambos orgánulos se reproducen en el interior celular por simple división, como las bacterias
4. Poseen doble membrana, siendo la externa aquella con la que la célula hospedadora rodeó a la célula simbionte que entró.
5. La membrana interna de las mitocondrias posee un tipo de fosfolípido exclusivo de las membranas de los organismos procariotas.
.
22. PRÁCTICAS




Microscopio
Huevos
Construcción microscopio casero
Funciones celulares
23. TAREAS









24. VÍDEOS
ANIMACIONES
7. LA CÉLULA
En 1665, el naturalista inglés Robert Hooke observó una fina lámina de corcho con un microscopio muy básico y distinguió en ella unas celdillas o huecos a las que denominó células. Sin embargo, realmente estaba observando las paredes celulares de células muertas, cuyo interior estaba vacío.
Poco después, en 1674, el holandés Antón van Leeuwenhoek construyó un microscopio rudimentario, de una sola lente, con el que observó gotas de agua, sangre, esperma, etc. Describió con gran detalle los seres y las células que observó, a los que denominó «animáculos».
Posteriores avances en el desarrollo del microscópico y diversas investigaciones llevaron a la conclusión de que las células no estaban vacías y que en su interior se podían observar estructuras.


De esta manera se establecieron:
7.1. Los principios de la teoría celular:
La teoría celular se debe a dos científicos alemanes, el botánico Mathias Schleiden y el zoólogo Theodor Schwann. En 1838, Schleiden señaló por primera vez que las plantas se componen de células. Al año siguiente, Schwann extendió esta generalización a los animales. La teoría celular no tardó en imponerse, pues agrupó un conjunto de datos que ya gozaban de consenso en la comunidad científica y desde entonces se acepta que la célula es la unidad básica de todos los organismos vivos.


En el año 1855, Rudolfh Virchow amplió la teoría celular y afirmó que las células solo surgen por división de otras células preexistentes, contradiciendo así la teoría (que aún entonces tenía muchos adeptos), de que las células pueden surgir por generación espontánea de la materia inanimada.
7.1. Los principios de la teoría celular:
En el año 1855, Rudolfh Virchow amplió la teoría celular y afirmó que las células solo surgen por división de otras células preexistentes, contradiciendo así la teoría (que aún entonces tenía muchos adeptos), de que las células pueden surgir por generación espontánea de la materia inanimada.
Durante el siglo XX, la teoría celular fue reafirmada y ampliada y es hoy uno de los conceptos unificadores más importantes de la biología. En su formulación actual, la teoría celular enuncia:
1) Los seres vivos están formados por células y productos celulares.
2) Las células se originan a partir de otras células.
3) Las reacciones químicas del organismo vivo tienen lugar dentro de células.
4) Las células contienen la información hereditaria de los organismos que integran y esta información se transmite de la célula madre a la célula hija.
Todas aquellas características que se hacen evidentes en un organismo complejo y nos permiten reconocerlo como un ser vivo, están presentes en cada una de las células que lo componen.
Las características de las células son:
- Tienen una organización compleja.
- Son sistemas abiertos: intercambian materia y energía con el medio.
- Realizan una serie de transformaciones químicas a las cuales se les da el nombre de metabolismo.
- Poseen un programa genético que guía el desarrollo de sus estructuras y su funcionamiento. Ese programa genético está inscripto en la estructura del ADN (ácido desoxirribonucleico) y contiene información para la síntesis de proteínas. Sin embargo, el ADN no participa en forma directa en la elaboración de proteínas. Para ello, la célula sintetiza una molécula intermediaria, el ARN (ácido ribonucleico), donde se transcribe la información genética almacenada en el ADN. El ARN es el artífice directo de la síntesis de proteínas, proceso también llamado traducción. Las proteínas son las ejecutoras del programa. Por lo tanto, la puesta en marcha de un programa genético requiere
- Tienen movimiento.
- Poseen receptores que les permiten captar señales del medio y responden a ellas.
- Se autorregulan.
- Se reproducen.
Tamaño celular
Las células miden típicamente unos pocos micrómetros (=1mm = 10-6m) de diámetro. Las células no sobrevivirían con volúmenes mayores. El límite al tamaño celular viene impuesto fundamentalmente por dos necesidades:Que la superficie celular, a través de la cual se realizan los intercambios con el medio, dé abasto para suministrarle a la célula los nutrientes necesarios y permitirle la eliminación de sus desechos. Cuanto mayor es el volumen de un cuerpo, proporcionalmente menor resulta su superficie. Tómese este simple ejemplo: un cubo A de 30 micrómetro de lado tiene una superficie de 900 micrómetros cuadrados y un volumen de 2700 micrómetro cúbico. Un cubo B de 10 micrómetros de lado tiene una superficie de 900 micrómetros cuadrados cada uno de los 27 cubos pequeños, lo que determina una superficie total de 27000 micrómetros cuadrados y un volumen de 1000 micrómetros cúbicos cada cubo pequeño para un total de 2700 micrómetros cúbicos. Si la superficie de estos cuerpos tuviera que ser utilizada para realizar intercambios, como ocurre con las células, el cuerpo B sería más eficiente que A, ya que dispone de una superficie relativamente mayor para su volumen.
Magnitud
| |||||||
Tamaños
|
Pesos
|
Sedimentación
| |||||
Metro
|
m
|
Gramo
|
g
|
Svedberg
|
(S)
| ||
Milímetro
|
mm
|
10 E-3 m
|
Miligramo
|
mg
|
10 E-3 g
| ||
Micrómetro o micra
|
µm
|
10 E-6 m
|
Microgramo
|
µm
|
10 E-6 m
| ||
Nanómetro
|
nm
|
10 E-9 m
|
Nanogramo
|
ng
|
10 E-9 m
| ||
Amgstron
|
A
|
10 E-10 m
|
Picogramo
|
pg
|
10 E-12 m
| ||
Unidades de peso molecular
|
Daltons
| 1g H= 6.023 E23 | |||||
Unidades de longitud utilizadas en Biología celular
- Tamaño celular: se expresa en micrómetros (millonésima parte del metro)
- Tamaño de estructuras subcelulares: se expresa en nanómetros (mil millonésima parte del metro)
- Tamaño de macromoléculas: se expresa en angstroms (diez mil millonésima parte del metro).

8. El Microscopio
Un microscopio óptico es un
microscopio basado en lentes ópticos. También se le conoce como microscopio de luz, (que utiliza luz o "fotones") o microscopio de campo claro. El desarrollo de este aparato suele asociarse con los trabajos de Anton van Leeuwenhoek. Los microscopios de Leeuwenhoek constaban de una única lente pequeña y convexa, montada sobre una plancha, con un mecanismo para sujetar el material que se iba a examinar (la muestra o espécimen). Este uso de una única lente convexa se conoce como microscopio simple, en el que se incluye la lupa, entre otros aparatos ópticos.
Un microscopio electrónico es aquél que utiliza electrones en lugar de fotones o luz visible para formar imágenes de objetos diminutos. Los microscopios electrónicos permiten alcanzar ampliaciones hasta 5100 veces más potentes que los mejores microscopios ópticos, debido a que la longitud de onda de los electrones es mucho menor que la de los fotones "visibles".
Un microscopio electrónico, como el de la imagen, funciona con un haz de electrones generados por un cañón electrónico, acelerados por un alto voltaje y focalizados por medio de lentes magnéticas (todo ello al alto vacío ya que los electrones son absorbidos por el aire). Un rayo de electrones atraviesa la muestra (debidamente deshidratada y en algunos casos recubierta de una fina capa metálica para resaltar su textura) y la amplificación se produce por un conjunto de lentes magnéticas que forman una imagen sobre una placa fotográfica o sobre una pantalla sensible al impacto de los electrones que transfiere la imagen formada a la pantalla de un ordenador. Los microscopios electrónicos producen imágenes sin ninguna clase de información de color, puesto que este es una propiedad de la luz y no hay una forma posible de reproducir este fenómeno mediante los electrones; sin embargo, es posible colorear las imágenes posteriormente, aplicando técnicas de retoque digital a través del ordenador.
9. TIPOS DE CÉLULAS
La célula. Es la estructura viva más sencilla que se conoce, es decir que es capaz de realizar las tres funciones vitales, que son nutrirse, relacionarse y reproducirse. Consta de dos partes que son la membrana plasmática y el citoplasma.
- Membrana plasmática. Es la capa que delimita la célula. Regula la entrada y salida de sustancias.
- Citoplasma. Es el contenido de la célula. En él se puede diferenciar un medio líquido denominadoplasma o citosol y una serie de estructuras denominadas orgánulos celulares. Los principales son los ribosomas, las vacuolas, las mitocondrias, el retículo endoplasmático, el aparato de Golgi y, sólo en las células que hacen la fotosíntesis, también los cloroplastos.
En el interior de la célula hay una o más moléculas de una sustancia denominada ADN. Se trata de unas moléculas muy alargadas, tan largas que tienen el aspecto de un hilo de coser, que contienen lainformación genética, es decir la información de como es y como funciona la célula. Una copia de estas moléculas se pasa a cada una de las células hijas para que puedan existir. Según que las moléculas de ADN estén dispersas en el citosol o rodeades de una membrana especial formando una estructura denominada núcleo, se diferencian dos tipos de células: las procariotas y las eucariotas.
ACTIVIDADES: 17 20 21 22 25 29 32 33 34 4 12 20 24 25 28 30 31 32 35 37 39 40 44 46 47 52 53 72
ACTIVIDADES: 17 20 21 22 25 29 32 33 34 4 12 20 24 25 28 30 31 32 35 37 39 40 44 46 47 52 53 72
ANIMACIONES
9.1. Células procariotas. Son las células que no tienen núcleo, es decir son las que presentan su ADN más o menos condensado en una región del citoplasma pero sin estar rodeado de una membrana. El ejemplo más importante de células procariotas son las bacterias. Son células muy sencillas, sus orgánulos prácticamente sólo son los ribosomas, los mesosomes (unos orgánulos exclusivos de estas células) y algunas también tienen unos flagelos muy sencillos.
- El ADN se encuentra libre y disperso por el citoplasma.
- No tienen orgánulos celulares como las mitocondrias, cloroplastos, aparato de Golgi, retículo, etc.
- Carecen de citoesqueleto y no tienen movilidad intracelular.
Existen tres dominios de seres vivos :
- Arqueobacterias - Archaea
- Eubacterias - Bacteria
- Eucariotas - Eukaria
Los Eucariotas son las células más diferenciadas y complejas.
Los otros dos dominios presentan células más sencillas : Organización procariota
Procariotas
|
Eucariotas
| ||
Bacteria
|
Archaea
|
Eukaria
| |
Adaptaciones genenerales
|
Sencillez
Rapidez y eficiencia metabólica Estrategia de la R |
Sencillez
Ambientes extremos |
Complejudad
Tamaño grande Estrategia de la K |
Tamaño
|
Pequeñas
Normalmente de 1 a 5 micras |
Pequeñas
Normalmente de 1 a 5 micras |
Grandes
Normalmente de 5 a 50 micras Entre 1.000 y 10.000 veces mayores que procariotastípicas |
Material genético
|
ADN circular
Sin nucleosomas: Sin histonas 1 Cromosoma |
ADN circular
Con nucleosomas: Con proteínas semejantes a histonas 1 Cromosoma |
ADN lineal
Con nucleosomas y estructuras superiores Ligado a histonas y otras proteínas Varios cromosomas |
Membranas internas
|
Pocas o ninguna
Sin membrana nuclear |
Ninguna
Sin membrana nuclear |
Muchas membranas internas
- Retículo endoplasmático - Golgi - Lisosomas - Vacuolas - Membrana nuclear |
Formadas por fosfolípidos
|
Formadas por éteres de terpenos
|
Formadas por fosfolípidos
| |
Pared celular
|
Casi siempre presente
Formada por peptidoglucano y otros compuestos |
Casi siempre presente
No formada por peptidoglucano |
Frecuente
Formada por polisacáridos (celulosa, quitina...) y otras sustancias |
Otros orgánulos
|
Ribosomas pequeños 70s
|
Ribosomas pequeños 70s (?)
|
Ribosomas grandes 80s
Mitocondrias y Plastos Microtúbulos |
Formas
|
No muy variadas
- Cocos - Bacilos - Espirilos - Filamentos |
No muy variadas
- Cocos - Bacilos - Filamentos - Aplanadas |
Muy variadas
|
Reproducción y
Sexualidad |
Reproducción asexual
Pueden tener procesos parasexuales |
Reproducción asexual
Pueden tener procesos parasexuales |
Reproducción sexual o asexual
- Asexual: Mitosis - Sexula: Meiosis y fecundación |
Metabolismo
|
Muy variado
|
Variado
|
Poco variado.
Todos aerobios |
Seres vivos: Células
| ||
- Dominio Bacteria :Bacterias y cianobacterias
| ||
- Dominio Archaea
| ||
- Dominio Eucaria
| ||
- Protistas : Varios reinos de algas y protozoos
| ||
- Reino Hongos
| ||
- Reino Animales
| ||
- Reino Plantas
| ||
Formas acelulares con información en Ácidos Nucleicos
| ||
- Virus : Genoma de ADN o ARN con faseextracelular: Virón
| ||
- Plásmidos, viroides, trasposones : Genoma de ADN o ARN sin fase extracelular
| ||
Elementos infecciosos sin Ácido nucléico
| ||
Priones: proteínas autotransformables de neuronas
| ||
ANIMACIONES
ACTIVIDADES: 10 1 2 3 5 6 8 8 35 36
La célula eucariota. Es puede definir como una estructura biológica constituida por tres partes denominadas membrana plasmática, citoplasma y núcleo , y que es capaz de realizar las tres funciones vitales. La célula eucariota es la unidad estructural y funcional de todos los organismos pluricelulares. Presenta formas y tamaños muy diferentes. Generalmente tienen una medida de unos unos 0,020 mm, pero algunas células eucariotas, como la yema del huevo de gallina, tienen más de un centímetro de diámetro
9.2.1. Células animales o heterótrofas: Se caracterizan por no presentar membrana de secreción o, si la presentan, nunca es de celulosa, por tener vacuolas muy pequeñas, por la carencia de cloroplastos y por presentar centrosoma, un orgánulo relacionado con la presencia de cilios y de flagelos.

9.2.2. Células vegetales o autótrofas: Se caracterizan por presentar una pared gruesa de celulosa situada en el exterior (sobre la membrana plasmática), por tener grandes vacuolas y cloroplastos (unos orgánulos de color verde debido a que contienden clorofila, que es la sustancia gracias a la cual pueden realizar la fotosíntesis) y por que no tienen ni cilios ni flagelos.
Las principales son:
|
10. ORGÁNULOS CELULARES Y SU FUNCIÓN
Las envolturas celulares son capas que separan el medio interno del exterior. Son: la membrana plasmática, que poseen todas las células (eucariotas y procariotas) y las membranas de secreción (que pueden faltar). Son membranas de secreción: la matriz extracelular (de animales), la pared vegetal (de vegetales) y la pared bacteriana (en bacterias).
11.1. La membrana plasmática
La membrana plasmática constituye el límite entre el citoplasma y el medio en el que se encuentra la célula y entre los orgánulos celulares y el citosol (hialoplasma), de manera que las biomembranas dividen al interior de la célula en numerosos compartimentos. De esta manera se explica que las membranas puedan constituir
el 60 % del citoplasma celular.Posee un espesor de 75 Ǻ (ángstrom). Al microscopio electrónico se presenta como una triple capa. Dos bandas oscuras externas de 20 Ǻ separadas por una interna de color claro de 35 Ǻ. En ocasiones, las láminas externas más oscuras pueden tener espesores diferentes.
Cualquier tipo de célula presenta este tipo de membrana de tres capas. Debido a su carácter generalizado se le denomina unidad de membrana (o membrana unidad). También se encuentra formando la envoltura exterior de muchos orgánulos como los cloroplastos, mitocondrias o vacuolas.
11.1.1. Composición química
Las membranas biológicas son conjuntos laminares constituidos aproximadamente por un 40 % de lípidos y un 60 % de proteínas principalmente; asociados a los lípidos y las proteínas también se encuentran oligosacáridos.
1. Lípidos. Son esencialmente anfipáticos, o sea que sus moléculas poseen un polo hidrófilo y un polo hidrófobo. Los más abundantes (en los glóbulos rojo humano) son los fosfolípidos (55 % del total de los lípidos), el colesterol (25%) y otros lípidos, glucolípidos y ácidos grasos(20 %) que son enteramente hidrófobos.
Los lípidos, cuando se encuentran en un medio acuoso, se disponen formando una doble capa, la bicapa lipídica, con las zonas polares (hidrófilas) orientadas hacia el exterior, y las zonas apolares (hidrófobas) protegiéndose mutuamente.
La bicapa lipídica no es una estructura rígida, sino que sus componentes se mueven en ella con libertad (Fig. 2), confiriéndole fluidez. Las moléculas de lípidos, más móviles, pueden girar sobre sí mismas (rotación) o intercambiar su posición con otras moléculas de la misma monocapa (difusión lateral). Es poco frecuente el intercambio entre moléculas situadas en monocapas distintas (flip-flop).▪ El colesterol contribuye grandemente a la fluidez de la bicapa, debido a que con su pequeño tamaño dificulta el establecimiento de interacciones hidrofóbicas entre las colas apolares de los lípidos de membrana.
2. Las proteínas se disponen intercaladas o adosadas a la bicapa de lípidos, y son de diverso tamaño y naturaleza.
Por su afinidad a los lípidos de la membrana hay dos tipos:
- Proteínas integrales o intrínsecas. Están internamente asociadas a los lípidos y son difíciles de separar de la bicapa. Constituyen aproximadamente el 70% de las proteínas de membrana y son insolubles en agua (hidrófobas)
- Proteínas periféricas o extrínsecas. Están débilmente asociadas a los lípidos, se separan con facilidad; son hidrosolubles y representan el 30% restante.
- Proteínas transmembranarias: atraviesan completamente la membrana.
- Proteínas de hemimembrana: su posición sólo abarca la mitad de la bicapa.
- Proteínas adosadas: colocadas por fuera de la bicapa, tanto hacia el interior como hacia el exterior de la célula y unidas a proteínas transmembrana o a lípidos
11.1.2. Estructura de la membrana plasmática
El modelo estructural de membrana más aceptado en la actualidad es el propuesto por Singer y Nicholson y que recibe el nombre de modelo del mosaico fluido. Según éste, todas las membranas celulares responden a un esquema arquitectónico común constituido, básicamente, por una bicapa lipídica a la que se unen los otros componentes de las membranas, es decir: proteínas y glúcidos.
La disposición en bicapa de los lípidos se debe al carácter bipolar que poseen estos constituyentes de la membrana
(fosfolípidos, colesterol, glicolípidos, etc.), en un medio acuoso, estos lípidos se disponen enfrentando sus partes hidrófobas dejando sus cabezas hidrófilas en contacto con el medio acuoso intra y extracelular.
El modelo estructural de membrana más aceptado en la actualidad es el propuesto por Singer y Nicholson y que recibe el nombre de modelo del mosaico fluido. Según éste, todas las membranas celulares responden a un esquema arquitectónico común constituido, básicamente, por una bicapa lipídica a la que se unen los otros componentes de las membranas, es decir: proteínas y glúcidos.
La disposición en bicapa de los lípidos se debe al carácter bipolar que poseen estos constituyentes de la membrana
(fosfolípidos, colesterol, glicolípidos, etc.), en un medio acuoso, estos lípidos se disponen enfrentando sus partes hidrófobas dejando sus cabezas hidrófilas en contacto con el medio acuoso intra y extracelular.
La situación de las proteínas (integrales o periféricas) en la membrana la determina su afinidad por el agua o los lípidos. Los azúcares (oligosacáridos), asociados a proteínas (glucopropteínas) o lípidos (glucolípidos), se sitúan en la cara extracelular de la membrana. A esta cubierta se la llama glucocáliz o cubierta celular.
Según lo expuesto, el modelo de mosaico fluido de membrana sostiene:
▪ Los lípidos y proteínas que forman la membrana plasmática constituyen un mosaico molecular. Están dispuestas unas junto a otras como las piezas de un mosaico.
▪ Los lípidos y proteínas pueden desplazarse en el plano de la bicapa lipídica: las membranas son fluidas.
▪ Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares. La asimetría se debe a que la presencia de oligosacáridos está restringida a la superficie de la cara externa y, además, la distribución de los lípidos en una y otra monocapa no es simétrica.



● Frontera física entre dos medios. Esto permite no sólo la separación del interior de la célula con respecto al medio exterior (intra y extracelular), sino también la formación de compartimentos en el interior de la célula eucariótica.
● Facilita que ocurran, de manera simultánea, pero sin mezclarse, una gran diversidad de reacciones químicas en sus diferentes orgánulos.
● La bicapa lipídica es una eficaz barrera para evitar el paso de sustancias hidrófilas (se evita la pérdida de sustancias intracelulares).
● Asegura el intercambio y transferencia de sustancias e información con el exterior y con otras células. La naturaleza lipídica de la membrana determina el tipo de sustancias que pueden atravesarla y, además, las proteínas que la forman, pueden intervenir de manera activa facilitando o impidiendo el transporte de esas sustancias.
Otras funciones de las diferentes membranas, determinadas por la presencia de proteínas específicas, son:
● Factores de reconocimiento celular. Conforman la “identidad antigénica” de cada individuo debido a que las proteínas específicas de la membrana celular constituyen una combinación única en cada individuo, que permite ser reconocida por las defensas inmunitarias.
● Receptores hormonales y de otras informaciones. Control del flujo de información entre las células y el medio, reciben la información que llega del medio, gracia a la existencia de a) receptores específicos de neurotransmisores y de hormonas, y b) el potencial de membrana responsable de la sensibilidad celular.
● Desempeñar funciones especiales gracias a las diferenciaciones que presentan algunas: invaginaciones (aumento de la superficie de intercambio), desmosomas (zonas de unión con otras células), etc.
11.1.3.1. Transporte a través de la membrana plasmática.
Las membranas son barreras de permeabilidad muy selectiva. Los mecanismos que utilizan las células para permitir el paso de sustancias varían en función de que se trate de moléculas pequeñas, que puedan atravesarla, o de moléculas más grandes, que deban ser englobadas y posteriormente liberadas por la propia membrana:
1. Moléculas pequeñas. La permeabilidad de las membranas celulares es altamente selectiva. El paso a su través de moléculas e iones está controlado por mecanismos de transporte específicos. Existen dos mecanismos básicos de transporte: el activo y el pasivo.
● Transporte pasivo: se realiza sin consumo de energía y a favor de un gradiente, ya que la sustancia pasa debido a que hay una diferencia de concentración o de carga eléctrica (o de ambos, electroquímico).
● Transporte activo: se realiza en contra de un gradiente (es decir, de la zona más diluida a la más concentrada), se requieren también proteínas transportadoras específicas y un aporte de energía (para realizar el “bombeo”), que se traduce en un consumo de ATP.
- Difusión simple: las sustancias solubles (apolares) pueden atravesar la membrana disueltas en la bicapa lipídica. El agua (y pequeñas moléculas arrastradas por ellas) atraviesa la membrana por ósmosis, lo hace a través de canales acuosos formados por proteínas transmembranarias, desde el lado de menor concentración salina al de mayor.
- Difusión facilitada: las sustancias polares (azúcares, aminoácidos), para pasar requieren la presencia de proteínas transportadoras (permeasas), a las que se une de manera específica la molécula a transportar y son liberadas de nuevo en el otro lado de la membrana.
La difusión simple y la facilitada se realizan a favor de un gradiente (de concentración o químico, eléctrico, o electroquímico) con lo que no requiere aporte de energía para realizarse.
En la difusión simple no intervienen proteínas mediando el paso de sustancias.
La difusión facilitada y el transporte activo se realizan con la mediación de moléculas de proteínas de la membrana.
En la difusión facilitada intervienen las proteínas transportadoras llamadas “permeasas” que, a pesar de su nombre, no realizan ninguna función enzimática. Las permeasas se unen de forma específica a la molécula que ha de ser transportada y por un cambio de conformación facilitan su paso
En la difusión facilitada intervienen las proteínas transportadoras llamadas “permeasas” que, a pesar de su nombre, no realizan ninguna función enzimática. Las permeasas se unen de forma específica a la molécula que ha de ser transportada y por un cambio de conformación facilitan su paso
Un ejemplo es la bomba de sodio-potasio, que mantiene el potencial electroquímico a ambos lados de la membrana de las células animales (bombea Na+ hacia el exterior de la célula y K+ hacia el interior).
2. Macromoléculas. La célula dispone de mecanismos que permiten incorporar o expulsar compuestos de mayor tamaño, por medio de deformaciones de la membrana.



- Endocitosis: incorpora partículas mediante una invaginación de la membrana en la que quedan incluidas. Luego, la invaginación se estrangula y forma una vesícula en el interior. Se distinguen dos tipos:
- Pinocitosis. Cuando el material captado es líquido (partículas disueltas).
- Fagocitosis. Capta partículas sólidas de mayor tamaño, y se forman vacuolas digestivas. Sólo la realizan células especializadas (por ejemplo, leucocitos).
- Exocitosis. Proceso opuesto al anterior. Se expulsan sustancias contenidas en una vesícula, al unirse ésta a la membrana plasmática y abrirse al exterior.
- Trancitosis. Proceso por el que las sustancias que penetran en la célula por endocitosis tras recorrer la totalidad del citoplasma salen al exterior por un proceso de exocitosis. Ejemplo: transporte de nutrientes por las células de las vellosidades intestinales.
ANIMACIONES








CUESTIONES: 1
11.1.4. Diferenciaciones de la membrana plasmática
Dependiendo de la función que la célula desempeñe, su membrana plasmática puede presentar diferentes especializaciones.
I.-Microvellosidades. Son evaginaciones que aumentan la superficie de intercambio. Por ejemplo, en las células intestinales, en las que las moléculas digeridas en el interior del tubo digestivo deben pasar al torrente circulatorio.


II.- Invaginaciones. Profundos entrantes con finalidad semejante. Por ejemplo, en las células de los túbulos contorneados de las nefronas (riñón), para la reabsorción de líquido y sales.
CUESTIONES: 1
11.1.4. Diferenciaciones de la membrana plasmática
Dependiendo de la función que la célula desempeñe, su membrana plasmática puede presentar diferentes especializaciones.
I.-Microvellosidades. Son evaginaciones que aumentan la superficie de intercambio. Por ejemplo, en las células intestinales, en las que las moléculas digeridas en el interior del tubo digestivo deben pasar al torrente circulatorio.
II.- Invaginaciones. Profundos entrantes con finalidad semejante. Por ejemplo, en las células de los túbulos contorneados de las nefronas (riñón), para la reabsorción de líquido y sales.
III.- Uniones intercelulares: para mantener adheridas y comunicadas células vecinas.
• Uniones occludens (impermeables). No dejan espacio entre las células e impiden el paso de sustancias, actuando a modo de barrera. Son frecuentes entre las células epiteliales.
• Uniones comunicantes. En ellas existe un pequeño espacio intercelular, con lo que las membranas no llegan a contactar y permite el paso de pequeñas moléculas entre dos células vecinas. Se pueden distinguir dos tipos:
Sinapsis, se realizan entre dos neuronas separadas por la hendidura sináptica.
Uniones en hendidura o de tipo gap (del ingles, hendidura), dejan entre sí una hendidura lo suficientemente ancha como para permitir el paso entre ellas de moléculas relativamente grandes.
• Uniones adherentes (desmosomas). El espacio intercelular aumenta y en la cara interna de la membrana plasmática se sitúa un material denso, denominado “placa”, hacia el que se dirigen haces de filamentos. Estas uniones se localizan en aquellos tejidos que se encuentran sometidos a esfuerzos mecánicos.
11.2. PARED CELULAR VEGETAL.
La pared celular es una forma especializada de matriz extracelular (segregada por la célula y excreta al exterior de la membrana plasmática), que se encuentra adosada a la membrana plasmática de las células vegetales, y que se caracteriza por su alto contenido encelulosa, lo que la hace ser gruesa, rígida y organizada.
11.2.1. Composición química de la pared vegetal
Como ya hemos señalado, esta formada principalmente por celulosa (homopolisacárido que se origina por la unión β (1→4) de la D-glucosa), pero también por: Hemicelulosa, Pectinas, Sales minerales y Agua
11.2.2. Estructura de la pared vegetal
Está constituida por tres capas, cada una con distinta composición y características. Desde fuera hacia dentro son:
I.- Lámina media: es la capa más externa y es común a las dos células adyacentes. Es delgada y flexible, y está compuesta principalmente por Pectinatos de calcio. Se encarga de mantener unidas las distintas células en los tejidos vegetales.II.- Pared primaria: capa relativamente delgada y semirrígida, típica de las células jóvenes, recién divididas (plantas en crecimiento). Está formada por celulosacon una abundante matriz hemicelulósica.
III.- Pared secundaria: capa muy gruesa formada por varias subcapas de celulosa, en cada una de las cuales las fibras de celulosa se disponen con distinta orientación, lo cual le da a la pared una gran rigidez y resistencia. La pared secundaria sólo se presenta en células maduras o ya muertas. Precisamente el grosor de la capa de celulosa hace que el citoplasma se vaya "asfixiando", y la célula acabe por morir.
11.2.3. Funciones de la pared vegetal
▪ Constituyen un exoesqueleto que protege a la célula, le da forma y le confiere resistencia, pero sin impedir su crecimiento.
▪ Es la responsable de que la planta se mantenga erguida.
▪ Impide que la célula se rompa, ya que interviene activamente en el mantenimiento de la presión osmótica intracelular.
▪ Permite la comunicación entre células adyacentes y con el exterior, para el intercambio de nutrientes y de información. Existen unos orificios que atraviesan la pared llamados punteaduras que se sitúan al mismo nivel en células vecinas. Estas punteaduras son atravesadas por puentes citoplasmáticos o plasmodesmos, que son prolongaciones del retículo endoplasmático.
11.3. MATRIZ EXTRACELULAR
Se encuentra por fuera de la membrana celular y desempeña un papel muy importante en la vida de la célula. Esta constituida por compuestos que segregan localmente la propia células.
El glucocálix (conjunto de cadenas de oligosacáridos) aparece en la cara externa de la membrana celular de muchas células animales. Tiene funciones de reconocimiento celular indispensables para la fecundación, reconocimiento de la célula a parasitar de virus y bacterias, adhesión de células para formación de tejidos y recepción de antígenos específicos para cada célula.
Su estructura consiste en una fina red de fibras de proteína inmersa en una estructura gelatinosa de glucoproteínas hidratadas, la sustancia fundamental amorfa.
En su composición química hay fundamentalmente: colágeno, elastina, fibronactina, glucoproteínas.
La función es primordialmente servir de unión y nexo en los tejidos conectivos, cartilaginoso y conjuntivo. Puede acumular sales, originando tejido óseo o quitina y dando lugar a exoesqueletos.
ANIMACIONES
CUESTIONES: 1 2 3 4 5 6 4
12. RETÍCULO ENDOPLASMÁTICO.
Está formado por un conjunto de membranas que limitan cavidades cerradas o cisternas de distintas formas: sáculos aplanados,vesículas globulares o tubos de aspecto sinuoso que delimitan un espacio interno, lumen o espacio cisternal contenido en el interior del retículo endoplásmatico y el espacio citosólico en el exterior del
retículo. Estas cavidades se comunican a menudo entre ellas y forman una red (retículo = pequeña red) característica de las células eucariotas.Sacos del retículo, por lo tanto, dobles membranas, separan el material nuclear del resto del citoplasma. Esta doble membrana está atravesada por una serie de poros que permiten el paso de grandes moléculas (ARN, subunidades ribosómicas, etc.).
Podemos distinguir dos tipos de retículo:
1.1. Retículo endoplasmático rugoso, que posee ribosomas adheridos a la cara de la membrana que da al hialoplasma (espacio citosólico). Se encuentra muy desarrollado en aquellas células que participan activamente en la síntesis de proteínas.
Dentro de sus funciones destacan: la síntesis de proteínas por parte de los ribosomas que pueden ser almacenadas en el del retículo donde maduran con frecuencia sufren una glucosilación de las proteínas (se unen a un glucido, glucoproteínas) y son transportadashacia otros orgánulos (aparato de Golgi, lisosomas), a la membrana celular o las propias membranas del retículo.
1.2. Retículo endoplasmático liso, constituido por finos túmulos o canalículos interconectados y cuyas membranas se continúan con las del REr, pero sin llevar adheridos ribosomas.
Sus funciones son: la síntesis lípidos de membrana (fosfolípidos, colesterol, etc.). Asimismo, en el retículo tiene lugar la detoxificación de aquellas sustancias perjudicialespara la célula producidas por su actividad vital o procedente del exterior (insecticidas, herbicidas, medicamentos, etc.). Las toxinas que proceden del medio externo llegan a nuestras células, son transformadas por este orgánulo en moléculas cuya toxicidad se ha visto reducida y que son de esta manera más fácilmente eliminadas.Gracia a la acción conjunta de ambos retículos queda completa la síntesis de los componentes básicos de toda membrana celular: proteínas y lípidos.
13. APARATO DE GOLGI.
El aparato de Golgi es un orgánulo membranoso que forma un sistema de cavidades aplanadas dispuestas ordenadamente. Cada una de estas cavidades recibe el nombre de sáculo. El conjunto de sáculos apilados recibe el nombre de dictiosoma. Además, pueden observarse toda una serie de vesículas más o menos esféricas a ambos lados y entre los sáculos relacionadas con el transporte de proteínas y lípidos desde y hacia el A. de Golgi.
El conjunto de todos los dictiosomas y vesículas celulares constituye el aparato o complejo de Golgi. Se encuentra más desarrollado cuanto mayor es la actividad celular y en las células secretoras.
El aparato de Golgi tiene dos caras distintas, la cara “cis”, “externa” o de formación y la cara “trans”, “interna” o de maduración.
Los dictiosomas no son estructuras celulares fijas, sino que se están renovando constantemente. Se forman a partir del retículo endoplásmatico. Este emite unas prolongaciones que se separan de él formando vesículas de transición (estas encierran en su interior el contenido de las cavidades del retículo, fundamentalmente proteínas, que se han fabricado en los ribosomas del retículo endoplasmático rugoso y cuyo destino es la secreción). Al unirse posteriormente dichas vesículas entre sí dan lugar a un sáculo (cara cis o de formación). Por otra parte, las cavidades golgianas opuestas (cara trans o de maduración) se fragmentan para formar vesículas cargadas de productos de secreción.
Funciones. El aparato de Golgi funciona como una planta “empaquetadora” y “distribuidora” de los productos fabricados por el R. E.● Embalaje y secreción. Las sustancias fabricadas en el R. E. para ser secretadas se incorporan a la cara “cis” o de formación del aparato de Golgi y se desplazan progresivamente hacia la cara “trans” o de maduración, desde donde se incorporan a las vesículas de secreción. Las vesículas se adosan a la membrana y por un proceso de exocitosis vierten su contenido al medio extracelular.
● Este proceso genera un reciclaje permanente de la membranacompensado por los mecanismos de endocitosis. Entre los productos secretados tienen especial interés los que forman la matriz extracelular que, en los vegetales, origina la pared celular.
● No todos los productos empaquetados por el aparato de Golgi tienen como destino el exterior de la célula. Por ejemplo, algunas vesículas, que contienen abundantes enzimas hidrolíticos, se transforman en lisosomas y permanecen en el citoplasma celular.
● En los retículos endoplasmáticos se sintetizan los componentes moleculares de las membranas (lípidos en el liso y proteínas en el rugoso) y el contenido de sus cavidades. Del retículo se desprenden vesículas que se incorporan a los sacos del aparato de Golgi, donde sus componentes moleculares experimentan modificaciones, glucolisaciones, por la adición de oligosacáridos, que se unen a proteínas (glucolisación de proteínas: glucoproteínas de membrana, anticuerpos) y a lípidos (glucolisación de lípidos: glucolípidos de membrana).
ANIMACIONES
14. LISOSOMAS.
Los lisosomas son orgánulos celulares, globulares que presentan formas muy diversas. Se originan a partir del retículo endoplásmico o del aparato de Golgi, de la misma manera que se originan las vesículas de secreción.
Constituyen el aparato digestivo de la célula. Contienen enzimas hidrolíticas (fosfatasa ácida, proteasa,...) que catalizan la hidrólisis o digestión de las macromoléculas; la membrana que limita a cada lisosoma impide que la célula sea digerida por estos enzimas líticos.
Se encuentran en las células eucariotas tanto animales como vegetales, la estructura de estos orgánulos es muy distinta de un tipo celular a otro e incluso en una misma célula. Este polimorfismo hace que no sea posible caracterizar a los lisosomas bajo criterios estrictamente estructurales y tan sólo la demostración de actividades líticas permite poner de manifiesto la naturaleza lisosomica de una inclusión celular.A pesar de esta diversidad anatómica, los lisosomas se clasifican en dos categorías:
Lisosomas primarios, recién formados a partir del Ap. de Golgi o del R.E., de forma ovalada o redondeada, que no han intervenido en ningún proceso de digestión y sólo contienen enzimas hidrolíticos.
Lisosomas secundarios, de forma variable (depende de la naturaleza de los sustratos y del estado de digestión en el que se encuentren), están implicados en algún proceso de digestión. Se forman a partir de la unión de los lisosomas primarios con endosomas que contienen sustratos procedentes del medio externo (vacuolas heterofágicas o digestivas) o interno (vacuolas autofágicas).
Funciones.
A.- Digestión celular
■ Vacuolas heterofágicas o digestivas (heterofagia) con sustratos procedentes del medio externo, cuya finalidad es alimenticia y defensiva.
Las partículas procedentes del medio externo penetran en la célula, mediante un proceso de invaginación de la membrana celular, que se separa de ésta, formándose una vacuola alimenticia o vacuola de endocitosis. Los lisosomas primarios se unen a esta vacuola, dando lugar a un orgánulo único, la vacuola digestiva o lisosoma secundario, que contiene no sólo enzimas hidrolíticos, sino también moléculas que va a digerir, por lo que se considera a esta vacuola el estómago celular.
Estas moléculas, una vez que son digeridas, pasan al citoplasma celular. Sin embargo, no todas las sustancias son digeridas y por tanto no pueden ser aprovechadas por la célula. A estas partículas no digeridas se las llama productos residuales, siendo expulsados al exterior de la célula después de que la vacuola digestiva se ha transformado en vacuola fecal.
■ Vacuolas autofagicas (autofagia) si el sustrato lo constituyen componentes de la propia célula. Este mecanismo interviene en los procesos de desarrolla y asegura la nutrición en condiciones desfavorables.
Los lisosomas producen la hidrólisis de moléculas u orgánulos de la propia célula, tal es el envejecimiento celular, estados de ayuno en los que la digestión de sus constituyentes celulares propician los alimentos que la células necesitan. Cuando esto ocurre, el proceso recibe el nombre de autofagia, que significa comerse a sí mismo.
B. Digestón extracelular. En algunas ocasiones, los lisosomas vierten su contenido al exterior de la célula, donde tiene lugar la digestión (digestión extracelular).
15. PEROXISOMAS.
Son orgánulos presentes en casi todas las células eucariotas. Al igual que los lisosomas, son compartimentos vesiculares que contienen enzimas, pero éstas, a diferencia de las de los lisisomas, intervienen en reacciones oxidativas en las que se consume gran cantidad de oxígeno.
RH2 + O2 ▬▬▬▬▬► R + H2O2
En esta reacción, RH2 representa una molécula orgánica que se oxida. Como resultado de este tipo de oxidaciones se forma como producto H2O2, peróxido de hidrógeno (agua oxigenada) , auténtico veneno para la célula; sin embargo, éste no afecta a la célula, ya que los peroxisomas contienen una elevada cantidad de la enzima catalasa que cataliza la reacción:
2 H2O2 ▬ catalasa ▬▬▬▬▬►2 H2O + O2
De esta manera desaparece el peróxido de hidrógeno en el mismo orgánulo en el que se ha producido, no afectando
al citosol que le rodea. Estos orgánulos se forman por gemación de la membrana del retículo endoplasmático y suelen localizarse junto a él.
16. GLIOXISOMAS.
Orgánulos exclusivos de las células vegetales. Son parecidos a los peroxisomas, y se encargan, durante la germinación, de transformar los lípidos almacenados en la semilla en glúcidos necesarios para el crecimiento del embrión hasta que la plántula pueda realizar la fotosíntesis. Poseen enzimas del ciclo del ácido glioxílico que es una variante del ciclo de Krebs de las mitocondrias que permite sintetizar azúcares a partir de grasas. Es indispensable en semillas en germinación.
SISTEMA DE ENDOMEMBRANAS
Es el conjunto de membranas citoplasmáticas internas: fundamentalmente RE, dictiosomas, membrana plasmática y vacuolas, y constituye una unidad funcional.
Son estructuras móviles, continuamente cambiantes, con el RE como fuente de crecimiento de las mismas. No participan en los plástidos ni las mitocondrias.
ANIMACIONES
17. VACUOLAS
Almacenamiento de sustancias de reserva. Vesículas de forma globular y de tamaño variable, destinadas a almacenar todo tipo de sustancias.
Se forman a partir de muchos tipos de orgánulos membranosos:
- R.E. (vesículas de transición, de autofagia...)
- Aparato de Golgi (Vesículas de secreción)
- Membrana plasmática (por endocitosis, vacuolas heterofágicas)
La estructura de las vacuolas es muy sencilla: consiste solamente en un territorio celular rodeado de una membrana. Al conjunto de vacuolas de una célula se denomina Vacuoma.
Las células vegetales presentan vacuolas de gran tamaño, que a veces desplazan a la periferia todo el citoplasma celular y el núcleo, y son escasas en número. Esta vacuola recibe el nombre de vacuola central. En cambio, las células animales poseen numerosas vacuolas de pequeño tamaño, habitualmente denominadas vesículas.
Funciones.
• Almacenar sustancias de reserva:
- Lípidos - Ácidos grasos
- Sales minerales - Proteínas solubles
- Proteínas precipitadas (granos de aleurona)
• Servir de vehículo de transporte de sustancias entre orgánulos citoplasmáticos (vesículas de transición, de secreción, etc.) y entre el exterior e interior de la célula (vacuolas o vesículas de endocitosis).
• Regulan la presión osmótica de la célula. Las vacuolas pueden llenarse o vaciarse de agua para mantener constante la concentración del citoplasma.
• Vacuolas pulsátiles, es un caso especial son las que eliminan el exceso de agua citoplasmática en células de organismos dulceacuícolas (que son hipertónicos respecto al medio).
CUESTIONES: 1 2
18. MITOCONDRIAS.
Son orgánulos presentes en el citoplasma de todas las células eucarióticas (aeróbicas). Su forma se asemeja a un cilindro alargado, difíciles de observar al microscopio óptico, al que aparecen como palitos o bastoncitos alargados. Lo más frecuente es que las mitocondrias estén dispersas en el hialoplasma, y su número depende del tamaño de la célula: unas pocas en las levaduras y de 1000 a2000 en una célula hepática. Son las encargadas de la obtención de la energía mediante la respiración celular, proceso de oxidación en el que intervienen las ATP sintetasas. La energía obtenida se guarda en forma de ATP.
Ultraestructura.
La observación al microscopio electrónico pone de manifiesto que cada mitocondria está separada del hialoplasma por una membrana continua de 60 Å (anstrom) de espesor, que es la membrana mitocondrial externa. Esta membrana externa está rodeada interiormente por una segunda membrana, igualmente continua y de 60 Å de espesor, que es la membrana mitocondrial interna. La membrana interna forma repliegues orientados hacia el interior de la mitocondria y son las llamadas crestas mitocondriales (por lo que su superficie es 5 veces mayor que la de la membrana externa). Las dos membranas mitocondriales delimitan dos compartimentos diferentes: un primer compartimento situado entre la membrana mitocondrial externa y la interna que es el espacio intermembrana; un segundo compartimento limitado por la membrana mitocondrial interna que es la matriz mitocondrial. En la matriz encontramos ADN y ribosomas mitocondriales.
- Membrana mitocondrial externa: Delimita completamente a la mitocondria. Su estructura es la misma que la del resto de las membranas celulares (una doble capa lipídica y proteínas asociadas), es muy permeable debido a que posee un gran número de proteínas que forma “canales” a través de los que pasan gran cantidad de moléculas.
- Espacio Intermembrana: De composición similar al hialoplasma por la permeabilidad de la membrana externa. Las enzimas que posee le permiten transferir la energía del ATP fabricado en la mitocondria a otros nucleótidos, por ejemplo el AMP
AMP + ATP ▬► 2 ADP
Las moléculas de ADP así formadas pueden atravesar la membrana interna y ser fosforiladas en moléculas de ATP.- Membrana mitocondrial interna: Posee gran superficie (unas 5 veces más que la externa) debido a los repliegues que forman las crestas. Es impermeable a gran cantidad de sustancias (iones,...). Contiene un 20% de lípidos y un 80% de proteínas, las cuales se pueden clasificar en tres grupos:
- Proteínas transportadoras que regulan el paso de metabolitos a través de la membrana interna (cadena transportadora de electrones).
- Enzimas de la cadena respiratoria, que catalizan las reacciones de oxidorreducción.
- Complejo enzimático. La ATP-sintetasa, cataliza la producción de ATP en la matriz, poseen tres subunidades:
- Esfera F1 o partícula elemental (90 Å de Ø) pegadas a la cara matricial de la membrana interna. Es la parte catalítica del complejo.
- Pedúnculo F0, que une las esferas a la membrana.
- Base hidrófoba integrada en la membrana mitocondrial interna.
- Matriz: Espacio interno que contiene numerosos iones y moléculas solubles, en particular una gran variedad de enzimas para llevar a cabo las rutas metabólicas que tienen lugar en su interior. Además, contiene ADN mitocondrial, ribosomas (mitorribosomas), ARNt.
- ADN mitocondrial (ADNmt), es doble y circular, lleva información genética que codifica para la síntesis de algunas proteínas.
- Ribosomas 70 S (mitorribosomas), libres o asociados a la membrana mitocondrial interna.
- Iones calcio y fosfato (Ca++, H2PO4-,...), ADP, ATP, Coenzima A,...
- Enzimas que intervienen en:
- Duplicación, transcripción y traducción del ADN mitocondrial.
- Respiración celular.
1º. Oxidaciones respiratorias: Cuya finalidad es obtener energía para la célula a partir de materia orgánica. Consisten en la degradación completa de moléculas orgánicas a CO2 para liberar energía y formar ATP. Tienen lugar en dos series de procesos, el ciclo de Krebs, la beta oxidación de los ácidos grasos y la cadena de transporte de electrones, que suceden, respectivamente, en la matriz y en la membrana mitocondrial interna.
2º. Producción de metabolitos precursoras para la biosíntesis de macromoléculas, se sintetizan en el hialoplasma.
3º. Síntesis de proteínas mitocondriales, ya que, poseen la maquinaria y la información necesaria para ello.
19. PLASTOS.



Los Cloroplastos están englobados en los Plastos o Plastidios, conjunto de orgánulos exclusivos de células vegetales que tienen un origen común. Poseen, al igual que las mitocondrias, información genética propia, ribosomas y una doble membrana. Hay tres tipos de plastos, aunque su estructura es muy semejante:
● Leucoplastos: (leuco = blanco). En partes no verdes del vegetal - las subterráneas, por ejemplo- entre ellos destacan losamiloplastos que acumulan almidón en los tejidos de reservas.
● Cromoplastos: (cromo = color). Dan color rojo, anaranjado o amarillo a muchos frutos, flores y otros órganos. Acumulan pigmentos fotosintéticos secundarios (xantofilas y carotenos) y reservas nutritivas.
● Cloroplastos: (cloro = verde). Son de color verde debido a la Clorofila que contienen. Se encargan de realizar la Fotosíntesis, captando energía luminosa.
19.1. Cloroplastos
Los cloroplastos se localizan en células vegetales fotosintéticas y, en los vegetales superiores, tienen forma lenticular (de lenteja). Su número suele ser de unos 40 por célula. Son de color verde debido a su elevado contenido en clorofila.
Ultraestructura
La observación al microscopio electrónico de un cloroplasto de una célula de vegetal superior nos revela que cada cloroplasto está rodeado de una doble membrana (externa e interna), la membrana externa separa el cloroplasto del hialoplasma, y la membrana interna, que delimita un estroma. Entre ambas existe un espacio, el espacio intermembrana. La Membrana Interna Cloroplástica(M.I.C.), a diferencia de lo que ocurre en las mitocondrias, carece de crestas y encierra un gran espacio central, el estroma (que contiene ribosomas, enzimas, ADN y ARNt , gránulos de almidón y gotas de lípidos), en el que se bañan un tercer tipo de membrana, lamembrana tilacoidal, que constituyen las paredes de unas pequeñas vesículas discoidales aplanadas (a modo de largos sacos cerrados), los tilacoides. En las membranas de los tilacoides se encuentran los pigmentos fotosintéticos, fundamentalmente clorofila y carotenoides. Los tilacoides están comunicados entre sí y encierran un tercer compartimento el espacio tilacoidal (o intratilacoidal). Existen unos tilacoides más alargados que a veces comunican con la M.I.C. y que son llamados lamelas o "tilacoides del estroma".
Los tilacoides no se distribuyen de manera uniforme por el estroma y en ocasiones forman pequeños grupos apilados (a modo de pilas de monedas) llamados grana (plural de grano en latín) debido al aspecto que presentan al observarlos con el microscopio. Normalmente los tilacoides se disponen paralelamente al eje mayor del cloroplasto.
En el estroma se encuentran también ribosomas (plastorribosomas) y moléculas de ADN, dobles y circulares, que poseen información para sintetizar algunas proteínas del cloroplasto.


La observación al microscopio electrónico de un cloroplasto de una célula de vegetal superior nos revela que cada cloroplasto está rodeado de una doble membrana (externa e interna), la membrana externa separa el cloroplasto del hialoplasma, y la membrana interna, que delimita un estroma. Entre ambas existe un espacio, el espacio intermembrana. La Membrana Interna Cloroplástica(M.I.C.), a diferencia de lo que ocurre en las mitocondrias, carece de crestas y encierra un gran espacio central, el estroma (que contiene ribosomas, enzimas, ADN y ARNt , gránulos de almidón y gotas de lípidos), en el que se bañan un tercer tipo de membrana, lamembrana tilacoidal, que constituyen las paredes de unas pequeñas vesículas discoidales aplanadas (a modo de largos sacos cerrados), los tilacoides. En las membranas de los tilacoides se encuentran los pigmentos fotosintéticos, fundamentalmente clorofila y carotenoides. Los tilacoides están comunicados entre sí y encierran un tercer compartimento el espacio tilacoidal (o intratilacoidal). Existen unos tilacoides más alargados que a veces comunican con la M.I.C. y que son llamados lamelas o "tilacoides del estroma".
Los tilacoides no se distribuyen de manera uniforme por el estroma y en ocasiones forman pequeños grupos apilados (a modo de pilas de monedas) llamados grana (plural de grano en latín) debido al aspecto que presentan al observarlos con el microscopio. Normalmente los tilacoides se disponen paralelamente al eje mayor del cloroplasto.
En el estroma se encuentran también ribosomas (plastorribosomas) y moléculas de ADN, dobles y circulares, que poseen información para sintetizar algunas proteínas del cloroplasto.
Funciones de los cloroplastos.Una de ellas es la síntesis de proteínas cloroplásticas, ya que, como hemos visto, poseen la maquinaria y la información necesaria para ello.
Pero la su principal función es llevar a cabo la Fotosíntesis, que consiste en la formación de materia orgánica a partir de compuestos inorgánicos (H2O, CO2, NO3=, SO4=, PO43-...), que reducen gracias a la energía captada por los pigmentos fotosintéticos.
En los tilacoides se encuentran los pigmentos captadores de luz, y la cadena transportadora de electrones (o cadena fotosintética), donde el ADP sé fosforila y pasa a ATP. Al ser la luz, digamos, el motor de este proceso, se le da el nombre de fotofosforilación. En los tilacoides se produce también la reducción del NADP+, que pasa a NADPH + H+. Ambos procesos constituyen lo que se conoce como fase lumínica de la fotosíntesis.
La energía acumulada, tanto en el ATP como en el NADPH + H+, será utilizada en la fase oscura para la fijación del CO2 a moléculas orgánicas. Esta fase oscura de la fotosíntesis tiene lugar en el estroma del cloroplasto.
Como ocurre en la mitocondria, en los cloroplastos se encuentran también esferas de ATP-sintetasa (o ATPasa), situadas en lamembrana interna del cloroplasto que mira hacia el estroma y en la membrana de los tilacoides. La ATPasa cataliza la reacción de formación de ATP
.
SEMEJANZAS Y DIFERENCIAS ENTRE
MITOCONDRIAS Y CLOROPLASTOS
| |||
Semejanzas
mitocondria/cloroplasto
|
Diferencias
| ||
Mitocondria
|
Cloroplasto
| ||
Estructura
|
Poseen una doble membrana, espacio interior (matriz/estoma), ribosomas 70 S, ADN circular y doble
|
La membrana mitocondrial interna presenta unos repliegues hacia la matriz denominadas crestas.
|
El cloroplasto posee un tercer tipo de membrana en su interior (las membranas tilacoidales) que delimitan un espacio llamado intratilacoidal.
|
Función
|
En ambos orgánulos tiene lugar untransporte de electrones en la membrana interna y, asimismo,formación de ATP por flujo de protones a través de los complejos enzimáticos ATP-sintetasa, de la membrana interna
|
La principal función de la mitocondria es larespiración celular. Es un proceso catabólico. En él se obtiene energía química útil para la célula (ATP) mediante la oxidación completa de la materia orgánica. Los electrones procedentes de estas oxidaciones son, en último término, transportados hasta el oxígeno molecular que se reduce a agua.
|
La principal función del cloroplasto es lafotosíntesis: Es un proceso anabólico (nutrición autótrofa). Su objetivo es pues sintetizar materia orgánica a partir de materia inorgánica. La energía para el proceso la obtienen de la luz solar y los electrones del agua, cuya rotura (fotolisis) libera O2
|



Recordemos que los organismos vivos más antiguos sobre la Tierra eran células procarióticas (3.500 millones de años), habiendo aparecido mucho más tarde las células eucarióticas (700 m.a. tienen los restos más antiguos encontrados de estos organismos). Por tanto, lo único que se puede afirmar es que probablemente, las células eucariotas evolucionaron a partir de procariotas entre los 3.500 y los700 m.a.
Actualmente existen dos teorías que tratan de explicar el posible origen de la célula eucariota.
La Teoría endógena propone que la célula eucariótica es el resultado del aumento de tamaño de la procariótica, junto a una progresiva diferenciación interna, cuyo resultado ha sido el alto grado de complejidad que posee.
La Teoría endosimbiótica, propuesta por la bióloga Lynn Margulis, considera que el alto grado de complejidad se debe a asociaciones entre células que en principio eran independientes entre sí. Este tipo de asociación debió consistir en una simbiosis, relación en la que ambas especies asociadas resultan beneficiadas.
En esta asociación las premitocondrias, que serían bacterias aerobias, conseguirían la oxidación de los alimentos, y las células hospedadoras obtendrían una ganancia en energía. A su vez, los precloroplastos, antiguas cianobacterias, suministrarían alimentos a las células hospedadoras mediante fotosíntesis, y obtendrían de ellas moléculas simples necesarias para realizar dicha función. La asociación de procariotas del tipo de espiroquetas daría lugar, por otro lado, a centríolos, cilios y flagelos.
Después de millones de años de dependencia mutua con las células hospedadoras, estos procariotas simbiontes perderían su independencia, conservando únicamente algunos vestigios de su forma de vida anterior.
De acuerdo con esta hipótesis, las antiguas bacterias habrían entrado al interior de la célula hospedadora por endocitosis, encerradas en unas vesículas endocíticas; esta circunstancia explicaría la doble membrana que poseen tanto mitocondrias como cloroplastos.
Esta teoría endosimbiótica se ve avalada por los siguientes hechos:
1. Tanto mitocondrias como cloroplastos contienen ADN del tipo de células procariotas, no asociado a proteínas histónicas.
2. Los dos orgánulos poseen ribosomas con los que sintetizan sus propias proteínas, también de tipo procariótico (70 S).
3. Ambos orgánulos se reproducen en el interior celular por simple división, como las bacterias
4. Poseen doble membrana, siendo la externa aquella con la que la célula hospedadora rodeó a la célula simbionte que entró.
5. La membrana interna de las mitocondrias posee un tipo de fosfolípido exclusivo de las membranas de los organismos procariotas.







































{kind=link}
21. IDEAS FUNDAMENTALES
|
|
22. PRÁCTICAS
Microscopio
Huevos
Construcción microscopio casero
Funciones celulares
Mitosis
Prácticas citología 2
Prácticas citología 2
Antibiograma
Células mucosa bucal 2
Extracción del ADN
Cultivo de bacterias
Conjunto de prácticas
Epidermis de cebolla 3
Epidermis de lirio
Mitosis en raíces de cebolla
Células en mitosis 2
Reproducción celular
Yogurt
Ósmosis en epidermis de cebolla
Sangre
Observación de células sanguíneas 2
Grupos sanguíneos
Células sanguíneas
Grupos sanguíneos 2
Frotis de sangre
Frotis de sangre 2
Determinación del grupo sanguíneo
Análisis de sangre
Células mucosa bucal 2
Extracción del ADN
Cultivo de bacterias
Conjunto de prácticas
Epidermis de cebolla 3
Epidermis de lirio
Mitosis en raíces de cebolla
Células en mitosis 2
Reproducción celular
Yogurt
Ósmosis en epidermis de cebolla
Sangre
Observación de células sanguíneas 2
Grupos sanguíneos
Células sanguíneas
Grupos sanguíneos 2
Frotis de sangre
Frotis de sangre 2
Determinación del grupo sanguíneo
Análisis de sangre
23. TAREAS
24. VÍDEOS
Cromosoma
Mapa cromosómico
Descomprimir el ADN
Formación del cromosoma 1
Formación del cromosoma 2
Código genético para fabricar proteínas
Transcripción y traducción. Dogma central de la biología
Transcripción 1
Transcripción 2
25. OTRAS PRESENTACIONESMapa cromosómico
Descomprimir el ADN
Formación del cromosoma 1
Formación del cromosoma 2
Código genético para fabricar proteínas
Transcripción y traducción. Dogma central de la biología
Transcripción 1
Transcripción 2
Teoría celular
La célula 1
La célula 2
Unicelulares y pluricelulares
Microorganismo
Citosol y citoesqueleto
Mitocondrias y cloroplastos
Núcleo
La célula 1
La célula 2
Unicelulares y pluricelulares
Microorganismo
Citosol y citoesqueleto
Mitocondrias y cloroplastos
Núcleo
26. CUESTIONES
Célula 1
Célula 2
Célula 3
Célula 4
Dibujos células 2
Preguntas selectividad importantes
Preguntas selectividad célula 1
Preguntas selectividad célula 2
Preguntas selectividad célula 3
Preguntas selectividad célula 4
Dibujos de célula animal y vegetal
Dibujos para completar de células
Actividad de escalas
Actividad célula
Actividad célula
Microscopio óptico
Citología 1
Completar imágenes células
Citología 2
Citología 3
Test citología resuelto
Núcleo
Métodos de estudio de la célula
27. REPASO



CUESTIONES: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 15 16 18 19 21 25 33 34 36 38 45 46 47 48 49 57 62 69 70 2 3 6 2 6 9
28. ACTIVIDADES EVALUABLES







No hay comentarios:
Publicar un comentario