El núcleo interfásico. Cromatina. Formación de cromosomas. Estructura del cromosoma. Tipos de cromosomas.




1.
Explica brevemente los niveles de organización de la cromatina.
La cromatina está formada de ADN y proteínas de dos tipos:
- Histonas: son proteínas muy básicas a causa de la abundante presencia de aminoácidos básicos, como la arginina y lisina, cargados positivamente, que neutralizan las cargas negativas de los grupos fosfatos de los nucleótidos. Son proteínas de bajo peso molecular.
- No histonas: son las más abundantes y son las que realizan funciones tan importantes como la replicación, transcripción y regulación del ADN.
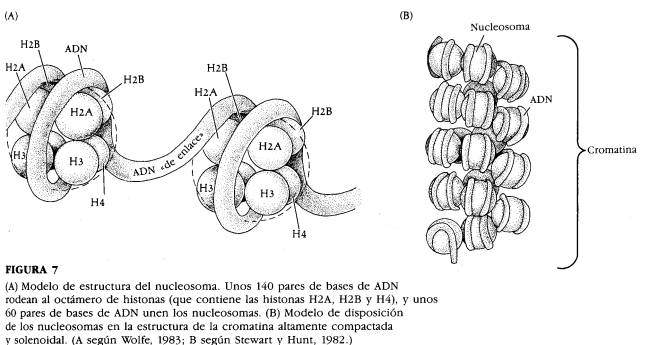
La doble hélice del ADN tiene un grosor de 2 nm y una longitud total (suma de las 23 moléculas diferentes de ADN características de nuestra especie) de 2,3 m. El primer nivel de condensación se le llama collar de perlas formada por una estructura repetida llamada nucleosoma de 10 nm de diámetro. Cada nucleosoma consta de un núcleo proteico formado por un octámero de histonas
2H2A, 2H2, 2H3, 2H4 y un filamento de la fibra de 2nm de ADN de 146 pares de nucleótidos, que forman casi dos vueltas a izquierda alrededor del nucleosoma.
Dos partículas de nucleosoma adyacentes, se unen mediante un segmento de ADN espaciador de longitud variable de unos organismos a otros. La quinta histona H1 se localiza fuera del nucleosoma y ocupa parte del ADN espaciador. Esta estructura es la básica en el estado de cromatina, también llamada nucleofilamento.
En el segundo nivel de espiralización el collar de perlas se compacta en una estructura en espiral de seis nucleosomas por vuelta llamado solenoide, de un diámetro de 30 nm. En esta estructura las seis histonas H1 se sitúan en el eje central de la hélice. Esta estructura acorta la longitud del collar de perlas cinco veces.
En el tercer grado de espiralización o dominio en forma de bucle, la fibra tiene un diámetro de 300 nm y forma bucles de entre 20.000 y 70.000 pares de bases de longitud. Su longitud se acorta unas 40 veces.
En el cuarto nivel la cromatina alcanza un diámetro de 700 nm y la longitud hasta ahora ha sido de unas 1000 veces.
El quinto y último grado de condensación es el de 1400 nm formando las cromátidas de los cromosomas. Su acortamiento es de unas 10.000 veces. Una molécula de ADN de 4 cm formará un cromosoma de 5,5 micras, lo que significa una reducción de longitud de unas 7000 veces.

- Histonas: son proteínas muy básicas a causa de la abundante presencia de aminoácidos básicos, como la arginina y lisina, cargados positivamente, que neutralizan las cargas negativas de los grupos fosfatos de los nucleótidos. Son proteínas de bajo peso molecular.
- No histonas: son las más abundantes y son las que realizan funciones tan importantes como la replicación, transcripción y regulación del ADN.
La doble hélice del ADN tiene un grosor de 2 nm y una longitud total (suma de las 23 moléculas diferentes de ADN características de nuestra especie) de 2,3 m. El primer nivel de condensación se le llama collar de perlas formada por una estructura repetida llamada nucleosoma de 10 nm de diámetro. Cada nucleosoma consta de un núcleo proteico formado por un octámero de histonas
2H2A, 2H2, 2H3, 2H4 y un filamento de la fibra de 2nm de ADN de 146 pares de nucleótidos, que forman casi dos vueltas a izquierda alrededor del nucleosoma.
Dos partículas de nucleosoma adyacentes, se unen mediante un segmento de ADN espaciador de longitud variable de unos organismos a otros. La quinta histona H1 se localiza fuera del nucleosoma y ocupa parte del ADN espaciador. Esta estructura es la básica en el estado de cromatina, también llamada nucleofilamento.
En el segundo nivel de espiralización el collar de perlas se compacta en una estructura en espiral de seis nucleosomas por vuelta llamado solenoide, de un diámetro de 30 nm. En esta estructura las seis histonas H1 se sitúan en el eje central de la hélice. Esta estructura acorta la longitud del collar de perlas cinco veces.
En el tercer grado de espiralización o dominio en forma de bucle, la fibra tiene un diámetro de 300 nm y forma bucles de entre 20.000 y 70.000 pares de bases de longitud. Su longitud se acorta unas 40 veces.
En el cuarto nivel la cromatina alcanza un diámetro de 700 nm y la longitud hasta ahora ha sido de unas 1000 veces.
El quinto y último grado de condensación es el de 1400 nm formando las cromátidas de los cromosomas. Su acortamiento es de unas 10.000 veces. Una molécula de ADN de 4 cm formará un cromosoma de 5,5 micras, lo que significa una reducción de longitud de unas 7000 veces.


2. Explica, sirviéndote de un esquema, la morfología del cromosoma metafásico.

El cromosoma representa la
máxima compactación de la cromatina. Gracias a esta compactación la longitud total de la molécula de ADN humana de 2,3 m, se reduce considerablemente haciendo posible el reparto equitativo del material genético.
Partes de un cromosoma:
Partes de un cromosoma:
- Centrómero: divide al cromosoma en dos partes iguales o no, llamados brazos.
- Cinetócoro: son los puntos situados a ambos lados del centrómero desde los cuales se polimerizan los microtúbulos que intervienen en la separación de las cromátidas de los cromosomas durante la etapa de anafase.
- Constricción secundaria: Estrechamiento que se sitúa cerca del telómero. Puede separar prociones de ADN llamadas satélites.
- Telómero: son estructuras protectoras (secuencias de ADN muy repetidas al final de las cadenas) que impiden que se pierda información, al no poderse completar totalmente la replicación de las cadenas de ADN. La desaparición de estas estructuras es prueba del envejecimiento celular.
- Satélites: porciones de ADN añadidas al cromosoma procedentes de otros cromosomas .
- Bandas: son segmentos de cromatina que se colorean de forma distinta mediante colorantes específicos y permiten un registro del patrón de bandas de cada cromosoma para su identificación y estudio.
- Dependiendo de la posición del centrómero hay cuatro tipos de cromosomas:

3. Explica la estructura y composición del núcleo interfásico.
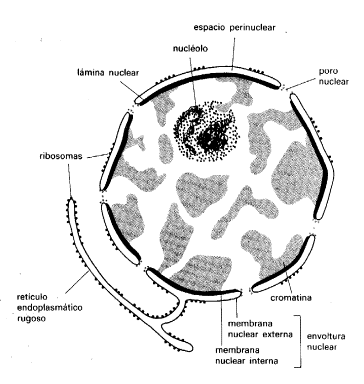
Estructuralmente el aspecto del núcleo depende de la fase del ciclo celular en que se encuentre la célula. Se habla de núcleo interfásico cuando la célula no está en división y mitótico cuando se observan los cromosomas.Los componentes del núcleo interfásico son:
- Envoltura nuclear: formada por una doble membrana con poros que controla y regula la comunicación entre el citoplasma y el nucleoplasma. La membrana externa es similar a la membrana plasmática con ribosomas adosados y se comunica con el retículo endoplasmático rugoso. La membrana interna presenta proteínas de membrana que sirven de anclaje a los componentes de la lámina nuclear (capa densa de proteínas fibrilares de características semejantes a los filamentos intermedios del citoesqueleto).
- Poros nucleares: orificios de 80 nm formados por el complejo del poro nuclear, estructura anular formada por dos grupos de 8 gránulos de ribonucleoproteínas. En su centro se sitúan ocho proteínas cónicas que dejan un canal de 10 nm que puede obturarse por una proteína central. Regulan el flujo de las subunidades ribosomales y pequeñas proteínas.
- Nucleoplasma o carioplasma: medio interno del núcleo. Dispersión coloidal en forma de gel con proteínas fibrilares para fijar el nucleolo y la cromatina.
- Nucleolo: de forma esférica sin membrana. Durante la división celular desaparece. Terminada la telofase y desespiralizados los cromosomas, se vuelve a formar. Está formado por ARN y proteínas. En él se sintetizan el ARN nucleolar que se convertirá en los diferentes tipos de ARNr constituyentes de los ribosomas. Su tamaño es mayor en células que realizan una elevada síntesis de proteínas y que por tanto, necesitan un elevado número de ribosomas.
- Cromatina: filamentos de ADN en distintos grados de condensación y proteínas. Forman ovillos cerca del nucleolo y la lámina media. La cromatina se forma a partir de los cromosomas descondensados cuando finaliza la división del núcleo. Se colorea mediante colorantes básicos. Puede presentarse en forma de heterocromatina (muy condensada y poco activa durante la interfase) y eucromatina (sin condensar y activa, es decir, puede ser transcrita).
4. ¿Cuál es la función del nucleolo?
La funció principal del nucleolo es la biosíntesis de ribosomas. En ellos se localizan las secuencias de ADN que codifican para los ARNr y las proteínas ribosomales. El ADN transcrito, ARN nucleolar, se fragmenta en trozos para formar los diferentes ARN ribosomales. Otros genes serán transcritos a ARNm que posteriormente serán traducidos para formar las proteínas ribosomales.
Los dos componentes ribosomales se acoplan en el nucleolo para formar las dos subunidades ribosomales por separado. Estas subunidades salen del núcleo atravesando la envoltura nuclear por los poros nucleares y llegan al citoplasma.
Cuando las dos subunidades se acoplan, el ribosoma se activa y traducirá los nuevos ARN mensajeros. En las células con una intensa síntesis de proteínas hay mayor cantidad de nucleolos para poder fabricar mayor número de ribosomas.


Los dos componentes ribosomales se acoplan en el nucleolo para formar las dos subunidades ribosomales por separado. Estas subunidades salen del núcleo atravesando la envoltura nuclear por los poros nucleares y llegan al citoplasma.
Cuando las dos subunidades se acoplan, el ribosoma se activa y traducirá los nuevos ARN mensajeros. En las células con una intensa síntesis de proteínas hay mayor cantidad de nucleolos para poder fabricar mayor número de ribosomas.

5. Define los siguientes términos:
i
i
Cromátida: La cromátida es cada una de las unidades longitudinales que forman un cromosoma. Es la molécula de ADN superenrollada (quinto grado de condensación y un grosor de 1400 nm) unida a su cromátida hermana (obtenida por replicación de la original que hace de molde).
Ambas cromátidas quedan unidas por el centrómero.
Ambas cromátidas quedan unidas por el centrómero.
Cromosoma: Representa la máxima compactación de
la cromatina. Esta alta compactación es la que permite el reparto equitativo del material genético a las células hijas durante la división celular. Los cromosomas están formados por diversos dominios estructurales en forma de bucles, estabilizados por un andamio de proteínas no histonas.
Centrómero: Divide el cromosoma en dos partes llamados brazos del mismo o diferente tamaño. Ocupa una posición variable, pero fija para cada uno de los cromosomas.
Los centrómeros contienen heterocromatina constitutiva, es decir, cromatina compactada y genéticamente inactiva en todas las células.
Según la posición del centrómero se clasifican los cromosomas en : metacéntricos, submetacéntricos, acrocéntricos y telocéntricos.


Las diferencias son: el cromosoma metafásico es el que se observa durante la metafase. Está formado por dos filamentos idénticos (cromátidas) que se disponen paralelos entre sí y están unidos por el centrómero. Una de las cromátidas será la molécula de ADN original del padre o madre que se utiliza como molde para fabricar la otra cromátida hija mediante el proceso de replicación.
El cromosoma anafásico es el que se observa durante la anafase. Está formado por un sólo filamento porque ya se han separado las cromátidas del cromosoma. Cada cromátida se dirige a uno de los polos de la célula para formar los nuevos núcleos.


Los centrómeros contienen heterocromatina constitutiva, es decir, cromatina compactada y genéticamente inactiva en todas las células.
Según la posición del centrómero se clasifican los cromosomas en : metacéntricos, submetacéntricos, acrocéntricos y telocéntricos.

6. Haz un esquema que represente la morfología del cromosoma metafásico. ¿Qué diferencias hay con el cromosoma anafásico?
El cromosoma anafásico es el que se observa durante la anafase. Está formado por un sólo filamento porque ya se han separado las cromátidas del cromosoma. Cada cromátida se dirige a uno de los polos de la célula para formar los nuevos núcleos.


7. Explica ayudándote de estos esquemas, los niveles de organización de la cromatina y señala las diferencias entre la cromatina interfásica y el cromosoma metafásico.

La cromatina está formada de ADN y proteínas de dos tipos:
- Histonas: son proteínas muy básicas a causa de la abundante presencia de aminoácidos básicos, como la arginina y lisina, cargados positivamente, que neutralizan las cargas negativas de los grupos fosfatos de los nucleótidos. Son proteínas de bajo peso molecular.
- No histonas: son las más abundantes y son las que realizan funciones tan importantes como la replicación, transcripción y regulación del ADN.
En la figura A se observa la doble hélice del ADN; tiene un grosor de 2 nm y una longitud total (suma de las 23 moléculas diferentes de ADN características de nuestra especie) de 2,3 m.
En la figura B se observa el primer nivel de condensación; se le llama collar de perlas formado por una estructura repetida llamada nucleosoma de 10 nm de diámetro. Cada nucleosoma consta de un núcleo proteico formado por un octámero de histonas 2 H2A, 2H2, 2H3, 2H4 y un filamento de la fibra de 2nm de ADN de 146 pares de nucleótidos, que forman casi dos vueltas a izquierda alrededor del nucleosoma. Dos partículas de nucleosoma adyacentes se unen mediante un segmento de ADN espaciador de longitud variable de unos organismos a otros.
La quinta histona H1 se localiza fuera del nucleosoma y ocupa parte del ADN espaciador. Esta estructura es la básica en el estado de cromatina, también llamada nucleofilamento.
La quinta histona H1 se localiza fuera del nucleosoma y ocupa parte del ADN espaciador. Esta estructura es la básica en el estado de cromatina, también llamada nucleofilamento.
En la figura C se observa el segundo nivel de espiralización; el collar de perlas se compacta en una estructura en espiral de seis nucleosomas por vuelta llamado solenoide, de un diámetro de 30 nm. En esta estructura las seis histonas H1 se sitúan en el eje central de la hélice. Esta estructura acorta la longitud del collar de perlas cinco veces.
En la figura D se observa el tercer grado de espiralización o dominio en forma de bucle. La fibra tiene un diámetro de 300 nm y forma bucles de entre 20.000 y 70.000 pares de bases de longitud. Su longitud se acorta unas 40 veces.
En la figura E se observa el cuarto nivel; la cromatina alcanza un diámetro de 700 nm y la longitud hasta ahora ha sido de unas 1000 veces.
En la figura E se observa el cuarto nivel; la cromatina alcanza un diámetro de 700 nm y la longitud hasta ahora ha sido de unas 1000 veces.
En la figura F, el quinto y último grado de condensación es el de 1400 nm formando las cromátidas de los cromosomas. Su acortamiento es de unas 10.000 veces. Una molécula de ADN de 4 cm formará un cromosoma de 5,5 micras, lo que significa una reducción de longitud de unas 7000 veces.
La cromatina interfásica es la eucromatina, cromatina poco compactada y activa puesto que puede ser transcrita. El grosor de la fibra de ADN es de 30 nm (collar de perlas). El cromosoma metafásico corresponde al ADN en su estado de máxima compactación, por tanto no presenta actividad y no puede ser transcrito. El grosor de la fibra es de 1400 nm.
8. Explica la relación que existe entre cromatina, cromosoma y cromátida.
La cromatina es un complejo de nucleoproteínas formado por la combinación del ADN con histonas, y es la sustancia que compone químicamente a los cromosomas. Cuando la cromatina se compacta por condensación en la Profase de la Mitosis o Meiosis da lugar a la formación de cuerpos visibles llamados cromosomas, entonces los términos cromatina y cromosomas se refieren a lo mismo, uno en estado amorfo ( cromatina) otro en estado compacto ( cromosomas).
Las cromátidas o brazos del cromosoma son uno de los elementos morfológicos que tienen en común todos los tipos de cromosomas. Cada cromátida está formada por sólo una molécula de ADN enrollada a la manera de espiral desde un extremo a otro.
9. Define los términos de: cromátida, cromosoma, centrómero y cinetocoro.
· Cromátida: La cromátida es una de las unidades longitudinales de un cromosoma duplicado, unida a su cromátida hermana (copia de la anterior) por el centrómero, es decir, la cromátida es una molécula de ADN enrollada y condensada.
· Cromosoma: cada uno de los pequeños cuerpos en forma de bastoncillos en que se organiza la cromatina del núcleo celular durante las divisiones celulares (mitosis y meiosis).
· Centrómero: es la construcción primaria. Es la zona por la que el cromosoma interacciona con las fibras del huso acromático desde la profase hasta la anafase, tanto en mitosis como en meiosis, y es responsable de realizar y regular los movimientos cromosómicos que tienen lugar durante estas fases. Divide el cromosoma en dos brazos. Dependiendo de su posición se tienen diferentes tipos de cromosomas: metacéntricos, submetacéntricos, acrocéntricos y telocéntricos.
·Cinetocoro: es una estructura proteica situada sobre los centrómeros de los cromosomas. Sobre esta estructura se anclan los microtúbulos del huso mitótico durante los procesos de división celular (meiosis y mitosis).
El cinetocoro está localizado en una zona específica del cromosoma, el centrómero.
10.Identifica las partes del cromosoma metafásico señaladas por los números.

OTRAS PREGUNTAS
2.- ¿Qué entiende por ciclo celular? ¿Qué significa “Omnis cellula ex cellula”?
El ciclo celular comprende el conjunto de modificaciones que experimenta una célula desde su formación hasta que completa su división.
La frase en latín alude a la división celular y corresponde al célebre aforismo del fisiólogo alemán Rudolph Virchow (cuyas aportaciones sirvieron para completar la teoría celular). Significa que toda célula proviene de otra.
3.- Exponga un comentario sobre el gráfico adjunto y aclare todos los aspectos que se reseñan.

Se trata de una representación esquemática del ciclo celular.
La interfase comprende las etapas G1, S y G2.
Fase G1 (del inglés gap, intervalo). Se sintetizan las proteínas necesarias para incrementar el tamaño celular. Su duración es muy variable y condiciona la de todo el ciclo.
Dentro del período G1 existe una situación límite superada la cual tiende a completarse el ciclo celular. Este límite se conoce como punto de restricción o punto R.
Algunas células entran en un estado modificado del período G1, llamado G0 (parada de ciclo). La permanencia en G0 es propia de células muy diferenciadas, como las neuronas o las fibras musculares estriadas.
Fase S. Tiene lugar la síntesis de ADN, esto es, su replicación. Además de ADN se sintetizan las histonas.
Fase G2. La célula suele aumentar ligeramente su tamaño y se prepara para la fase M. Los filamentos de cromatina empiezan a formar estructuras más compactas.
La fase M es la última etapa del ciclo y no hay procesos de biosíntesis. La división celular comprende la del núcleo (mitosis) y la del citoplasma (citocinesis).
4.- ¿Varía la cantidad de ADN durante el ciclo celular?
En la célula recién originada el contenido de ADN nuclear es el normal, es decir, el que corresponde a los cromosomas formados por una sola cromátida o molécula de ADN. Así se mantiene durante el período G1 de la interfase. Esta cantidad “normal” de ADN suele anotarse como 2C, 2x o 2n. Posteriormente tendrá lugar la fase S, que se inicia cuando el ADN nuclear empieza a replicarse y termina cuando éste finaliza su replicación.
Por consiguiente, tras la duplicación del ADN nuclear en la fase S, el contenido es el doble del normal, situación que se mantiene durante el período G2 de la interfase. Cada cromosoma está formado por dos cromátidas (hermanas). Este contenido de ADN se anota como 4C, 4x o 4n.
Durante la mitosis tiene lugar el reparto equitativo de los cromosomas, de forma que cuando finaliza la división el contenido de ADN nuclear en cada una de las dos células resultantes vuelve a ser el normal o propio del comienzo de la interfase.
5.- ¿Qué representa el gráfico adjunto? Identifique las partes numeradas

Se trata de los cambios en el contenido de ADN nuclear durante las correspondientes fases o etapas del ciclo celular.
Interfase = 1, 2 y 3. 1 = período G1. 2 = fase S. 3 = período G2 (aquí se observa que el contenido de ADN, tras la replicación, es el doble del inicial). 4 = fase M (división celular). 5 = etapa G1 del ciclo siguiente.
6.- Construya un gráfico para representar la cantidad de ADN nuclear durante un ciclo celular, sabiendo que: el tiempo de la interfase es 18 h (horas), el ADN se ha sintetizado durante 6 h y la fase de división, que comienza 5 h después, dura 2 horas.

Total interfase = 18 h (G1 = 7 h. Fase S = 6 h. G2 = 5 h).
Fase M = 2h.
Duración del ciclo celular: 20 horas.
7.- ¿A qué se llama interfase? Exponga los acontecimientos que tienen lugar.
Se llama interfase al conjunto de procesos que experimenta la célula recién originada hasta que comienza su propia división. Se distinguen tres etapas: G1, S y G2.
En el período G1 (del inglés gap, intervalo) se sintetizan las proteínas necesarias para que la célula aumente de tamaño. Comienza cuando termina la fase M y dura hasta que se inicia la replicación del ADN. Su duración es bastante variable.
El período S o de síntesis de ADN se caracteriza por la replicación o duplicación del ADN nuclear. En las células de los mamíferos, por lo común, dura unas 7 horas. Como resultado de la replicación cada cromosoma está formado por dos cromátidas hermanas, unidas mediante el centrómero.
La fase o período G2 tiene una duración más corta (alrededor de 3 horas en los mamíferos) y la célula puede aumentar ligeramente su tamaño. Se transcriben y traducen genes que codifican las proteínas necesarias para la división celular y, a menudo, se duplican los centriolos. El período G2 finaliza cuando empieza la condensación de los cromosomas para comenzar la fase M (división celular).
8.- ¿Tiene siempre la misma duración el ciclo celular? Cite factores que influyan en la duración del mismo.
En general, la duración del ciclo celular es muy variable.
En los organismos unicelulares la duración está condicionada sobre todo por la disponibilidad de nutrientes y la temperatura del medio.
En los organismos pluricelulares las células tienen ciclos de distinta duración. Mediante experimentos apropiados se ha llegado a demostrar que los tiempos del ciclo celular difieren incluso en células del mismo tejido.
Los factores que influyen en la duración del ciclo celular son: el tamaño celular, el contacto con otras células, la disponibilidad de nutrientes, la temperatura, la edad, etc.
Durante el ciclo celular existen unos puntos para controlar, principalmente, el tamaño de la célula y la replicación del ADN.
9.- ¿Cuál es la fase de duración más variable en el ciclo celular? ¿Qué significado tiene la fase G0? ¿A qué se llama fase M?
La fase de duración más variable es la primera parte de la interfase, esto es, el período G1. Las variaciones en la duración del período G1 son las que determinan que el ciclo celular sea corto o largo. En un organismo hay células que están en fase G1 unas horas, unos días e incluso años. Además, cuando una célula se detiene dentro de su ciclo, lo hace siempre en G1.
Algunas células entran en un estado modificado del período G1, llamado G0 (parada de ciclo). La permanencia en G0 es propia de aquellas células que experimentan un proceso de diferenciación muy notorio, como las neuronas o las fibras musculares estriadas. Estas células detienen su ciclo y permanecen en un estado de reposo aparente, ya que presentan actividad metabólica y son funcionales, pero al no sobrepasar el punto de restricción no prosiguen el ciclo ni llegarán a dividirse.
La fase M corresponde a la división celular y en ella tiene lugar la mitosis y la citocinesis. Suele durar alrededor de 1 ó 2 horas.
10.- En relación con el cromosoma y el ciclo celular, exponga una interpretación del gráfico adjunto.

Se han representado los diferentes estados del cromosoma a lo largo del ciclo celular. Se puede interpretar que los dibujos A y B corresponden a la interfase y los restantes a la fase M.
El dibujo C corresponde a la metafase, D y E a la anafase y F, a la telofase.
El dibujo A corresponde al período G1 de la interfase. En el B se observa que ha empezado la replicación o síntesis de ADN (fase S).
11.- Interprete esta otra versión del estado del cromosoma durante el ciclo celular. ¿Qué conclusión puede aportar?

Se puede interpretar que los dibujos A y B corresponden a la fase M y los otros tres, a la interfase.
Fase M (división celular): A = metafase. B = anafase.
Interfase: C = G1. D = fase S. E = G2.
Conclusión: el cromosoma está formado por una sola molécula de ADN, desde la anafase hasta el comienzo de la fase S, o por dos, desde el final de S hasta el inicio de la anafase.
Nota.- El dibujo E también se podría considerar como profase.
12.- ¿Cómo es el ciclo celular en las células cancerosas? En relación con la quimioterapia, ¿a qué se llama cronoterapia?
En las células cancerosas está alterado el control normal de la división. Estas células crecen y se dividen de modo incesante, invadiendo otros tejidos, pues no presentan el fenómeno de la inhibición por contacto (frenar la división al contactar con otras células). En los cultivos de células normales, pasado un cierto tiempo, tras realizar un número determinado de divisiones, toda la población celular deja de dividirse y el cultivo envejece y muere. Por el contrario, el cultivo de células cancerosas crece indefinidamente.
Ensayando con dos tipos de cáncer hematológico se ha descubierto que las células tumorales carecen de una molécula encargada de regular el llamado ritmo circadiano, esto es, nuestro reloj biológico interno que regula los ciclos de sueño y vigilia. La molécula del reloj biológico que está inactiva en el cáncer es BMAL1, encargada de controlar que ciertos genes se expresen de forma rítmica. Por esta razón las células cancerosas estarían permanentemente despiertas (activas).
Este descubrimiento podría tener implicaciones a la hora de diseñar estrategias de tratamiento para los pacientes, administrándoles la quimioterapia en el momento del día más adecuado para que las células malignas respondan mejor (cronoterapia).
Así, pues, la cronoterapia consiste en hallar el momento para optimizar la aplicación del tratamiento con quimioterapia.
13.- ¿A qué se llama “senescencia replicativa”?
Las células de mamífero no se dividen infinitamente sino un número limitado de veces antes de diferenciarse en células altamente especializadas. Por ejemplo, los fibroblastos humanos en un medio de cultivo estándar se dividen entre 25 y 50 veces.
Se ha observado que la proliferación celular va disminuyendo su velocidad y finalmente se detiene, fenómeno que se ha denominado “senescencia replicativa”.
14.- ¿Qué característica del ciclo celular presenta una mayor relevancia?
Una de las características esenciales del ciclo celular es la existencia de mecanismos de control para conocer la fiabilidad del proceso y bloquearlo en el caso de que no se hayan realizado adecuadamente todos los procesos. Estos puntos de control garantizan, principalmente, la replicación y reparación del ADN así como la entrada y salida de la mitosis.
15.- ¿Cuántos puntos de control suelen considerarse en el ciclo celular? ¿Qué aspectos son supervisados en las fases G1, G2 y M?
Se han postulado cuatro puntos en los que se controla el estado de la célula y del medio extracelular, con objeto de decidir si se realizan o restringen las acciones propias de cada una de las fases del ciclo. Estos cuatro puntos de control son: el de restricción (R) y, sobrepasado éste, otros tres ubicados al final de G1, de G2 y en la fase M.
El primer punto de control se encuentra en G1, justo después del punto de restricción. Aquí se controla el tamaño celular y si las condiciones del entorno son favorables.
Al final de G2 (2º punto de control) se controla si está replicado todo el ADN, si es favorable el entorno y si el tamaño celular es apropiado para la división.
El 3er punto de control se halla en la fase M, entre la metafase y anafase. Se controla principalmente si todos los cromosomas están unidos al huso mitótico, así como si están alineados en el ecuador celular. Si se detecta que uno de los cinetocoros no se encuentra unido a los microtúbulos, se genera una señal al sistema de control que bloquea la activación de proteínas implicadas en la separación de las cromátidas hermanas.
16.- ¿Es de carácter extracelular el control del ciclo celular?
El control del ciclo celular se presenta a dos niveles, intracelular y extracelular.
Las células en los organismos pluricelulares proliferan solo cuando se requieren más células. La entrada al ciclo celular no es un proceso autónomo de la célula, pues se requiere la presencia de señales adecuadas, esto es, factores solubles de naturaleza proteica (mitógenos), procedentes del medio extracelular o de otras células.
17.- ¿Cómo se ejerce a nivel intracelular el control del ciclo?
El control del ciclo a nivel intracelular se realiza previa fosforilación, mediante quinasas específicas, de unas proteínas clave para posibilitar la replicación, la mitosis y la citocinesis.
La actividad de estas quinasas, llamadas Cdk (Cyclin dependent kinases) depende de unas proteínas activadoras llamadas ciclinas, que se sintetizan en determinados momentos del ciclo celular y posteriormente se degradan (el número de ciclinas identificadas es distinto en eucariotas inferiores y superiores).

En resumen: la regulación a nivel intracelular es llevada a cabo por complejos Cdk-ciclinas y sus inhibidores, entre otras proteínas. El fundamento del control del ciclo celular radica esencialmente en los distintos complejos Cdk-ciclinas, que controlan el inicio de las fases S y M.
19.- ¿Qué relación tiene la proteína p27 con el ciclo celular?
La proteína p27 (masa molecular 27 kDa, esto es, 27000 u) se une a ciclinas y Cdk inactivando el complejo ciclina-cdk, circunstancia que bloquea la entrada en la fase S.

20.- ¿Qué relación existe entre la proteína p53 y el ciclo celular?
La proteína p53 (masa molecular 53000 u, o sea, 53 kDa) es un regulador de la proliferación de la mayoría de los tipos celulares. Se trata de un factor de transcripción que, al fijarse en una secuencia concreta del ADN, activa los genes implicados en el programa de apoptosis o en la detención del ciclo celular. Concretamente, la inhibición de la proteína p53 provoca una proliferación celular.
21.- ¿Cómo actúa la p53?
Cuando el ADN presenta un daño limitado, aumenta la concentración de proteína p53. Este aumento desencadena una serie de reacciones y estimula la síntesis de otra proteína, llamada p21, la cual, por su acción inhibidora, provoca la inactivación de un complejo ciclina-Cdk (Cdk2-Ciclina E) y la consiguiente detención del ciclo.
Esta situación proporciona tiempo para la actividad de los sistemas de reparación del ADN, pues mientras persista la inactivación de dicho complejo la célula no puede entrar en la fase S.
Cuando el ADN es reparado, la proteína p53 se libera de la región promotora del gen p21, provocando el descenso en los niveles de p21. Esto permite restaurar la actividad del complejo ciclina-Cdk y la continuación del ciclo.
Nota.- La p53 es una proteína integrada por 393 aminoácidos y, estructuralmente, presenta 3 dominios. Es inestable en condiciones normales porque se encuentra unida a otra proteína, que funciona como un “marcador”, para que la citada p53 se degrade. El incremento de la concentración de p53 requiere la previa separación de su “marcador”, llevada a cabo por otros mediadores.
En células normales, el nivel de la proteína p53 es bajo porque es ubiquitinada y destruida por Mdm2. Los daños del ADN y otras señales de estrés pueden hacer que no se una a Mdm2 e incrementar su concentración, activando la transcripción del gen p21.
22.- ¿Qué significado tiene la expresión “el guardián del genoma”?
Cuando el ADN sufre daño, el gen p53, que codifica a la proteína p53, se activa y el incremento de la misma desencadena la ralentización o detención del ciclo celular, dando tiempo para que actúe el sistema de reparación de ADN, razón por la cual se ha denominado “el guardián del genoma”.
El gen p53 se halla en el brazo corto del cromosoma 17 y activa su expresión en situación de estrés o daño celular. Bloquea la proliferación o división de la célula alterada e induce la apoptosis (suicidio celular).
Nota.- Cuando fue descubierto, en 1979, el gen p53 se consideró dentro de los oncogenes (genes que promueven el cáncer). Pero, en 1989, pasó a integrar el grupo de los supresores tumorales. Ello se debió a que su primera identificación se obtuvo de una de sus formas mutadas y no de la normal o natural.
23.- ¿Qué son los genes supresores de tumores?
Ciertas proteínas, llamadas inhibidores de Cdk, y algunos factores de transcripción, como la proteína p53, tienen la función de impedir la proliferación celular. Los genes que codifican estas proteínas se llamaron “genes supresores de tumores”, como por ejemplo, el gen p53.
La mutación de los genes que codifican dichas proteínas, o bien, la disfunción de las mismas, implica la pérdida de control del ciclo celular y la incapacidad para detenerlo, lo cual conduce a la proliferación celular (tumorogénesis).
Las células que presentan los dos alelos del gen p53 mutados, no tendrán proteína p53 funcional y, por consiguiente, continuarán dividiéndose a pesar del daño en su genoma, circunstancia que, en general, desencadenará un proceso canceroso.
24.- Enumere las bases conceptuales del ciclo celular.
Fueron las contribuciones de los científicos galardonados con el Premio Nobel de Medicina de 2001 las que sirvieron para establecer las bases conceptuales del ciclo celular, que podemos exponer en cinco puntos:
1) El ciclo celular puede considerarse como una secuencia de acontecimientos organizados temporalmente.
2) El inicio de cada uno de ellos es consecuencia de la finalización de los anteriores.
3) La relación entre unos acontecimientos y otros se establece directamente o a través de algún tipo de señales.
4) La finalización de una etapa antes de comenzar la siguiente conlleva la existencia de controles a lo largo del ciclo.
5) Ciertos acontecimientos actúan como limitantes para la progresión del ciclo
25.- Resuma las características del ciclo celular.
• El ciclo celular es un conjunto de procesos ordenados que lleva a cabo la célula que va a dividirse. Comprende la interfase y la división celular.
• El control del ciclo celular se presenta a dos niveles, intracelular y extracelular.
• El control intracelular está a cargo de mediadores proteicos que ejercen una acción activadora o inhibidora.
• Hay un punto de restricción y tres puntos de control, regulados por distintas combinaciones de Cdk-ciclinas.
• La entrada al ciclo celular requiere señales adecuadas (mitógenos), procedentes del medio extracelular o de otras células.
• Cuando una célula no es necesaria o se convierte en una posible amenaza por estar infectada o alterada, puede morir por necrosis o apoptosis.
2.- Defina el concepto de apoptosis.
Apoptosis se puede definir como la muerte fisiológica de las células.
Aunque dicho término se considera sinónimo de muerte celular programada, la desaparición de muchas células no está prefijada, sino que es consecuencia de cambios impredecibles en su entorno. La apoptosis es generalmente inducida por agentes externos a la célula, con frecuencia producidos por otras células del organismo. Son las señales de pervivencia, de naturaleza cambiante en distintos períodos, las que impiden el inicio del suicidio celular. La regla general es que el destino de la célula no sólo depende del tipo de señales que reciba en cada instante, sino también de las que haya recibido antes (una excepción a esta regla es la actuación de los linfocitos citotóxicos).
La expresión “muerte celular programada” es en cierto modo equívoca, pues sugiere que la célula tiene fijada genéticamente una fecha de muerte. Lo correcto es interpretar que la célula posee un programa de autodestrucción, pero que sólo será ejecutado ante señales o situaciones muy concretas. Por esta razón se ha propuesto la denominación de suicidio inducido.
Nota.- El término apoptosis deriva del griego arcaico y alude a la caída natural de las hojas. Fue acuñado en 1972 por John Kerr y colaboradores.
3.- ¿Qué se requiere para desencadenar la apoptosis? ¿En qué momento del ciclo celular se toma la decisión de iniciar la apoptosis?
Las células de un ser pluricelular deben estar coordinadas mediante señales químicas para optimizar su fisiología. La respuesta a estas señales es selectiva e implica la recepción de la molécula señal mediante una proteína receptora específica, seguida de la correspondiente transducción, que transforma la señal extracelular en señales intracelulares que modificarán su actividad.
En ausencia de señales concretas o en presencia de señales “de muerte” la célula pone en marcha un programa que provoca su propia destrucción, teniendo lugar una degradación progresiva de los materiales y estructuras celulares, proceso denominado apoptosis o muerte celular controlada.
Existen proteínas extracelulares que, como las neurotrofinas, pueden favorecer la supervivencia, o por el contrario inducir la apoptosis, como ocurre con el ligando de Fas.
La decisión de proseguir el ciclo o entrar en la vía terminal de apoptosis se toma en el punto de control situado al final de la fase G1del ciclo celular.
4.- Haga un resumen sobre las diversas señales que regulan el suicidio celular.
La célula recibe señales de multiplicación celular que promueven la expresión de los llamados protooncogenes, que además de iniciar tal multiplicación, intervienen en la activación de las ciclinas para garantizar el curso de todas las etapas del ciclo celular.
Señales provenientes del medio extracelular, tales como el factor de necrosis tumoral (TNF) y el ligando del receptor Fas, inducen el suicidio celular.
La iniciación del suicidio se realiza mediante activación de las proteasas ICE.
Cuando la célula recibe menos señales de supervivencia disminuye la expresión de los genes inhibidores del suicidio, como Bcl-2.
5.- ¿Cómo actúan los linfocitos citotóxicos (o verdugos)?
Los linfocitos citotóxicos o verdugos, al entrar en contacto con las células infectadas, llevan a cabo la secreción de unos gránulos citoplásmicos que contienen proteínas citotóxicas: perforina y granzimas (las dos más importantes son la granzima A y la granzima B).
La perforina interacciona con las membranas celulares y permite el acceso de las granzimas al interior de la célula diana, las cuales inducen muerte celular por apoptosis.
Las granzimas desempeñan un papel crucial, pues intervienen en la activación proteolítica de las enzimas ICE de la célula alterada o infectada, desencadenando el suicidio de la misma, sin que en este caso influyan las señales que la célula reciba de su entorno.
Nota.- Algunos autores concluyen que el sistema perforina-granzima B induce apoptosis a través de la activación de caspasas, implicando a la vía mitocondrial, y que el sistema perforina-granzima A induce apoptosis a través de la generación de radicales libres, también por la vía mitocondrial, pero en ausencia de activación de caspasas
6.- ¿A qué se refiere el llamado “carácter altruista” de la apoptosis?
Los seres pluricelulares recurren a la apoptosis en muchos procesos fisiológicos y patológicos. El carácter altruista de la apoptosis alude a que la célula se autodestruye en beneficio del organismo. De esta forma, la apoptosis, también llamada muerte celular controlada, contribuye al desarrollo y bienestar del organismo.
Este programa de muerte celular ocurre, por ejemplo, en células infectadas por virus (evitando con ello la producción de nuevas partículas víricas) o en las que tengan importantes alteraciones en su ADN.
7.- ¿Qué significado tiene la frase: “La muerte celular forma parte de la vida”?
En la ontogénesis de cada ser vivo pluricelular, la muerte de ciertas células es necesaria para el correcto desarrollo orgánico.Concretamente, un embrión humano de pocas semanas tiene los dedos unidos por una membrana cuyas células se eliminan a sí mismas, dejando los dedos bien separados.
Además, todo ser vivo debe desprenderse de las células que a lo largo de su vida han ido acumulando mutaciones y errores susceptibles de convertirlas en cancerosas. Cualquier desarreglo que afecte a la capacidad de morir de estas células puede tener consecuencias letales para el organismo.
8.- ¿A qué se refiere la frase: “La muerte es escultora de vida”?
A lo largo del desarrollo embrionario la muerte celular modela la forma del embrión eliminando tejidos, como si fuese un escultor. Así, por ejemplo, la cola y las branquias del renacuajo desaparecen durante la metamorfosis en rana.
En el caso humano, la cola del embrión también es eliminada gracias a la muerte celular, quedando esculpido un feto sin cola, y la formación de la mano, mediante eliminación del tejido interdigital, ya se ha mencionado en la cuestión anterior.
9.- ¿Es lo mismo necrosis celular que apoptosis? ¿Qué factores pueden causar necrosis?
La necrosis o muerte accidental se produce cuando la célula sufre daño grave. La célula lesionada se hincha y la membrana celular se rompe, expulsando al medio su contenido, situación que suele desencadenar un proceso inflamatorio y provocar un daño más o menos severo en la matriz intercelular y en las células vecinas.
Por el contrario, en la apoptosis no hay expulsión de materiales celulares y, por lo general, las células que mueren de esta forma son células sanas, esto es, potencialmente funcionales, a diferencia de las que mueren por necrosis.
En la necrosis las células pueden ser alteradas por una serie de factores: lesiones, hipoxia, toxinas, agentes infecciosos, etc.
10.- ¿En qué consiste la apoptosis o muerte celular programada (MCP) y cuáles son las características morfológicas?
Actualmente se consideran tres formas o modalidades de muerte celular programada (MCP): tipo 1, apoptosis, que es la más común; tipo 2, autofagia, mediante acción de los lisosomas; tipo 3, muerte vesicular no lisosomal.
La apoptosis es el tipo de MCP más frecuente y consiste, fundamentalmente, en la activación de las enzimas ejecutoras del programa. Se trata de una muerte “natural” y las células se autodestruyen en un período de tiempo que en general oscila alrededor de 60 minutos, pero sin expulsar al medio su contenido.
Morfológicamente se caracteriza por: retracción celular, disminución de la turgencia, desorganización del citoesqueleto y de la lámina nuclear, invaginación de la membrana nuclear, fragmentación del material cromosómico (la rotura del ADN en grandes fragmentos sería la señal irreversible que conduce a la apoptosis), formación de protuberancias en la superficie celular. Finalmente, la célula origina varias vesículas o cuerpos apoptóticos (con orgánulos y fragmentos nucleares), que en general son fagocitados por los macrófagos.
11.- ¿Qué relación hay entre la apoptosis y las mitocondrias?
Las mitocondrias son los orgánulos que producen la energía de la célula, por lo que cualquier disfunción conduce a la desregulación del medio intracelular. En tales circunstancias la mitocondria deja de producir eficientemente energía y libera una serie de moléculas que pueden desencadenar la apoptosis.
Una de estas moléculas es el citocromo c, proteína crucial en la transferencia de electrones propia de la respiración celular, pero que en el citosol activa la vía de las caspasas.
La mitocondria puede liberar también el factor inductor de la apoptosis (AIF), una proteína cuya actividad apoptótica pasa por su transporte al núcleo y es independiente, al menos en parte, de la vía de las caspasas.
15.- ¿Por qué se dice que el gen p53 puede desempeñar el papel de policía o de asesino?
Este gen vela por la integridad del genoma y obliga a la célula a interrumpir su multiplicación y a reparar su lesión genética cuando ésta se produce, es decir, detiene el ciclo celular, actuando como policía, hasta que se termina la reparación, razón por la cual fue denominado “guardián” del genoma.
Las mutaciones que provocan la disfunción del gen p53 tienen como consecuencia que las células alteradas puedan seguir multiplicándose y sufrir nuevas transformaciones genéticas favorables a la tumoración (en torno al 50 % de los casos de cáncer en humanos presentan mutaciones en dicho gen).
Por otra parte, el gen p53 participa en la apoptosis o muerte celular programada, conduciendo a las células al suicidio cuando el daño en el ADN es tan significativo que rebasa la capacidad de reparación. Así, pues, cuando la lesión en el ADN es demasiado importante, el gen p53 desempeña el papel de asesino puesto que desencadena la muerte de la célula.
Nota.- El gen p53 está localizado en el brazo corto del cromosoma 17 y codifica a la proteína p53, cuya concentración intracelular debe estar bien regulada, pues si el nivel es demasiado alto puede exacerbar la apoptosis. El regulador principal de p53 es Mdm2, que actúa “marcando” a p53 para que sea ubiquitinada y degradada.
La proteína p53 es capaz de unirse a lugares específicos del ADN, regulando la expresión de otros genes que controlan funciones celulares esenciales. Esta es la razón de la conexión del gen p53 con procesos tales como la carcinogénesis, el ciclo celular y la activación de la apoptosis.
16.- ¿Cuál es la probable relación entre una mutación temprana del gen p53 y el desarrollo del cáncer de pulmón?
La proteína p53 (codificada por el gen p53), que bloquea el ciclo celular cuando el ADN está alterado, puede llegar a inducir apoptosis en la célula cuyo ADN esté severamente dañado. La mutación temprana del gen p53 tenderá a impedir que células anormales mueran por apoptosis, dejando vía libre al proceso de tumorogénesis.
17.- Busque en Internet el artículo “Apoptosis, señalización celular y transformaciones tumorales en Drosophila” (Ginés Morata. Centro de Biología Molecular “Severo Ochoa”, CSIC-UAM, Madrid). Encuentre la respuesta a las siguientes cuestiones:
A) ¿Qué funciones cumple la apoptosis?
B) ¿Cómo se puede inducir apoptosis?
C) ¿Es posible mantener vivas durante mucho tiempo células apoptóticas?
A) La mortalidad celular programada, o apoptosis, cumple varias funciones muy importantes durante el desarrollo animal.
Es necesaria para muchos procesos morfogenéticos, así como para impedir que haya exceso de células o eliminar células dañadas o debilitadas durante el desarrollo.
La apoptosis ha de estar regulada de forma muy precisa porque su desregulación da lugar frecuentemente a procesos tumorales y su exceso está relacionado con la aparición de enfermedades neurodegenerativas en humanos.
B) Existen órganos, como el disco imaginal de ala de Drosophila, donde no hay apoptosis en el desarrollo normal. Sin embargo, este proceso se puede inducir mediante tratamientos de estrés con irradiación (rayos X) o choque térmico, siempre que se haya inhibido previamente la actividad caspasa.
En estas condiciones se ha observado que las células emiten las moléculas señalizadoras Decapentaplegic (Dpp) y Wingless (Wg), las cuales inducen proliferación adicional en las células vecinas. Dpp y Wg son los morfógenos principales en el desarrollo deDrosophila, y sus homólogos en vertebrados (TFG-ß/BMP y Wnt) tienen funciones similares.
En experimentos recientes se ha estudiado el papel específico de las señales Dpp y Wg en el proceso, descubriendo que el estímulo proliferativo se debe únicamente a Dpp mientras que Wg tiene una actividad inhibitoria destinada a regular el efecto de Dpp.
C) También se ha descubierto, en experimentos con Drosophila, que se pueden mantener vivas indefinidamente las células apoptóticas mediante el uso de un inhibidor general de la actividad caspasa, la proteína de baculovirus P35.
19.- ¿Qué genes regulan la muerte celular de los organismos más complejos que Caenorhabditis elegans?
Relacionados con la regulación de la muerte celular, los seres humanos poseen dos genes muy parecidos a los del nematodo, los llamados ICE y Bcl-2, cuyas familias se componen al menos de 6 miembros cada una.
La expresión de los genes de la familia ICE promueve la apoptosis.
En la otra familia hay genes antagonistas, pues Bcl-2 y Bcl-XL favorecen la supervivencia impidiendo la activación o los efectos de las proteasas ICE, mientras que, por el contrario, Bax, Bad, Bag y Bak promueven el suicidio, oponiéndose a los efectos de los otros dos de su propia familia y apoyando la actividad de las citadas proteasas.
20.- ¿Está de acuerdo con la existencia de una cierta especialización de los genes reguladores de la muerte celular?
Se ha descubierto que la supervivencia de una población celular concreta viene asegurada en momentos diferentes de su existencia por la expresión de distintos genes de la familia Bcl-2. La expresión de Bcl-XL es indispensable, por ejemplo, para la supervivencia de los linfocitos embrionarios, mientras que la de Bcl-2 lo es para la de los linfocitos adultos.
A esta especialización de los genes reguladores de la muerte celular se superpone la diversidad de las reacciones que presentan las células ante determinada señal. Puesto que una misma señal puede desencadenar la expresión de diferentes genes en poblaciones celulares distintas, provocará la muerte de unas y la supervivencia de otras, circunstancia que apunta hacia la enorme complejidad de la regulación genética y molecular del suicidio celular.
21.- ¿Cómo se comportan las células normales y las cancerosas cuando emigran a otros órganos?
Una célula normal que emigre a un órgano que no es el suyo tendrá serios problemas de supervivencia, puesto que allí no recibe las señales necesarias para impedir que actúe su programa de autodestrucción.
Por el contrario, una célula cancerosa, cuyo programa de suicidio está inutilizado, puede seguir viviendo en ausencia de las señales apropiadas, circunstancia que permite colonizar nuevos órganos y formar metástasis.
22.- ¿Hay apoptosis en los procesos de tumorogénesis?
Se dan todos los grados intermedios entre los tumores cuyas células mueren por apoptosis en número considerable, pero se multiplican aún más aprisa, y aquellos cuyas células se multiplican prácticamente sin freno apoptótico, que son los más nocivos y los que en el caso humano suelen tener un pronóstico infausto.
OTROS ENLACES CON PREGUNTAS RELACIONADAS:
8. Explica la relación que existe entre cromatina, cromosoma y cromátida.
La cromatina es un complejo de nucleoproteínas formado por la combinación del ADN con histonas, y es la sustancia que compone químicamente a los cromosomas. Cuando la cromatina se compacta por condensación en la Profase de la Mitosis o Meiosis da lugar a la formación de cuerpos visibles llamados cromosomas, entonces los términos cromatina y cromosomas se refieren a lo mismo, uno en estado amorfo ( cromatina) otro en estado compacto ( cromosomas).
Las cromátidas o brazos del cromosoma son uno de los elementos morfológicos que tienen en común todos los tipos de cromosomas. Cada cromátida está formada por sólo una molécula de ADN enrollada a la manera de espiral desde un extremo a otro.
9. Define los términos de: cromátida, cromosoma, centrómero y cinetocoro.
· Cromátida: La cromátida es una de las unidades longitudinales de un cromosoma duplicado, unida a su cromátida hermana (copia de la anterior) por el centrómero, es decir, la cromátida es una molécula de ADN enrollada y condensada.
· Cromosoma: cada uno de los pequeños cuerpos en forma de bastoncillos en que se organiza la cromatina del núcleo celular durante las divisiones celulares (mitosis y meiosis).
· Centrómero: es la construcción primaria. Es la zona por la que el cromosoma interacciona con las fibras del huso acromático desde la profase hasta la anafase, tanto en mitosis como en meiosis, y es responsable de realizar y regular los movimientos cromosómicos que tienen lugar durante estas fases. Divide el cromosoma en dos brazos. Dependiendo de su posición se tienen diferentes tipos de cromosomas: metacéntricos, submetacéntricos, acrocéntricos y telocéntricos.
·Cinetocoro: es una estructura proteica situada sobre los centrómeros de los cromosomas. Sobre esta estructura se anclan los microtúbulos del huso mitótico durante los procesos de división celular (meiosis y mitosis).
El cinetocoro está localizado en una zona específica del cromosoma, el centrómero.
10.Identifica las partes del cromosoma metafásico señaladas por los números.
{kind=link}

1- Telómero
2- Cinetocoro
3- Histonas H1
4- Nucleosomas
5- Constricción secundaria
6- Bandas
7- Centromero
11. Es idéntico el material genético de dos cromosomas homólogos. ¿y de dos cromátidas hermanas?. Razona la respuesta.
Entre cromosomas homólogos el material tiene el mismo número y tipo de genes aunque pueden contener diferente información (alelos), por tanto, tiene que ser diferente ya que
uno se hereda de la madre y otro del padre durante la fecundación.
En
cromátidas hermanas el material genético es idéntico a menos que haya
habido algún error durante la replicación de éste durante la fase S del
ciclo celular. Durante la mitosis una célula que tiene
cromosomas con dos cromátidas se divide para dar origen a una célula con
una sola cromátida.
OTRAS PREGUNTAS
2.- ¿Qué entiende por ciclo celular? ¿Qué significa “Omnis cellula ex cellula”?
El ciclo celular comprende el conjunto de modificaciones que experimenta una célula desde su formación hasta que completa su división.
La frase en latín alude a la división celular y corresponde al célebre aforismo del fisiólogo alemán Rudolph Virchow (cuyas aportaciones sirvieron para completar la teoría celular). Significa que toda célula proviene de otra.
3.- Exponga un comentario sobre el gráfico adjunto y aclare todos los aspectos que se reseñan.
Se trata de una representación esquemática del ciclo celular.
La interfase comprende las etapas G1, S y G2.
Fase G1 (del inglés gap, intervalo). Se sintetizan las proteínas necesarias para incrementar el tamaño celular. Su duración es muy variable y condiciona la de todo el ciclo.
Dentro del período G1 existe una situación límite superada la cual tiende a completarse el ciclo celular. Este límite se conoce como punto de restricción o punto R.
Algunas células entran en un estado modificado del período G1, llamado G0 (parada de ciclo). La permanencia en G0 es propia de células muy diferenciadas, como las neuronas o las fibras musculares estriadas.
Fase S. Tiene lugar la síntesis de ADN, esto es, su replicación. Además de ADN se sintetizan las histonas.
Fase G2. La célula suele aumentar ligeramente su tamaño y se prepara para la fase M. Los filamentos de cromatina empiezan a formar estructuras más compactas.
La fase M es la última etapa del ciclo y no hay procesos de biosíntesis. La división celular comprende la del núcleo (mitosis) y la del citoplasma (citocinesis).
4.- ¿Varía la cantidad de ADN durante el ciclo celular?
En la célula recién originada el contenido de ADN nuclear es el normal, es decir, el que corresponde a los cromosomas formados por una sola cromátida o molécula de ADN. Así se mantiene durante el período G1 de la interfase. Esta cantidad “normal” de ADN suele anotarse como 2C, 2x o 2n. Posteriormente tendrá lugar la fase S, que se inicia cuando el ADN nuclear empieza a replicarse y termina cuando éste finaliza su replicación.
Por consiguiente, tras la duplicación del ADN nuclear en la fase S, el contenido es el doble del normal, situación que se mantiene durante el período G2 de la interfase. Cada cromosoma está formado por dos cromátidas (hermanas). Este contenido de ADN se anota como 4C, 4x o 4n.
Durante la mitosis tiene lugar el reparto equitativo de los cromosomas, de forma que cuando finaliza la división el contenido de ADN nuclear en cada una de las dos células resultantes vuelve a ser el normal o propio del comienzo de la interfase.
5.- ¿Qué representa el gráfico adjunto? Identifique las partes numeradas
Se trata de los cambios en el contenido de ADN nuclear durante las correspondientes fases o etapas del ciclo celular.
Interfase = 1, 2 y 3. 1 = período G1. 2 = fase S. 3 = período G2 (aquí se observa que el contenido de ADN, tras la replicación, es el doble del inicial). 4 = fase M (división celular). 5 = etapa G1 del ciclo siguiente.
6.- Construya un gráfico para representar la cantidad de ADN nuclear durante un ciclo celular, sabiendo que: el tiempo de la interfase es 18 h (horas), el ADN se ha sintetizado durante 6 h y la fase de división, que comienza 5 h después, dura 2 horas.
Total interfase = 18 h (G1 = 7 h. Fase S = 6 h. G2 = 5 h).
Fase M = 2h.
Duración del ciclo celular: 20 horas.
7.- ¿A qué se llama interfase? Exponga los acontecimientos que tienen lugar.
Se llama interfase al conjunto de procesos que experimenta la célula recién originada hasta que comienza su propia división. Se distinguen tres etapas: G1, S y G2.
En el período G1 (del inglés gap, intervalo) se sintetizan las proteínas necesarias para que la célula aumente de tamaño. Comienza cuando termina la fase M y dura hasta que se inicia la replicación del ADN. Su duración es bastante variable.
El período S o de síntesis de ADN se caracteriza por la replicación o duplicación del ADN nuclear. En las células de los mamíferos, por lo común, dura unas 7 horas. Como resultado de la replicación cada cromosoma está formado por dos cromátidas hermanas, unidas mediante el centrómero.
La fase o período G2 tiene una duración más corta (alrededor de 3 horas en los mamíferos) y la célula puede aumentar ligeramente su tamaño. Se transcriben y traducen genes que codifican las proteínas necesarias para la división celular y, a menudo, se duplican los centriolos. El período G2 finaliza cuando empieza la condensación de los cromosomas para comenzar la fase M (división celular).
8.- ¿Tiene siempre la misma duración el ciclo celular? Cite factores que influyan en la duración del mismo.
En general, la duración del ciclo celular es muy variable.
En los organismos unicelulares la duración está condicionada sobre todo por la disponibilidad de nutrientes y la temperatura del medio.
En los organismos pluricelulares las células tienen ciclos de distinta duración. Mediante experimentos apropiados se ha llegado a demostrar que los tiempos del ciclo celular difieren incluso en células del mismo tejido.
Los factores que influyen en la duración del ciclo celular son: el tamaño celular, el contacto con otras células, la disponibilidad de nutrientes, la temperatura, la edad, etc.
Durante el ciclo celular existen unos puntos para controlar, principalmente, el tamaño de la célula y la replicación del ADN.
9.- ¿Cuál es la fase de duración más variable en el ciclo celular? ¿Qué significado tiene la fase G0? ¿A qué se llama fase M?
La fase de duración más variable es la primera parte de la interfase, esto es, el período G1. Las variaciones en la duración del período G1 son las que determinan que el ciclo celular sea corto o largo. En un organismo hay células que están en fase G1 unas horas, unos días e incluso años. Además, cuando una célula se detiene dentro de su ciclo, lo hace siempre en G1.
Algunas células entran en un estado modificado del período G1, llamado G0 (parada de ciclo). La permanencia en G0 es propia de aquellas células que experimentan un proceso de diferenciación muy notorio, como las neuronas o las fibras musculares estriadas. Estas células detienen su ciclo y permanecen en un estado de reposo aparente, ya que presentan actividad metabólica y son funcionales, pero al no sobrepasar el punto de restricción no prosiguen el ciclo ni llegarán a dividirse.
La fase M corresponde a la división celular y en ella tiene lugar la mitosis y la citocinesis. Suele durar alrededor de 1 ó 2 horas.
10.- En relación con el cromosoma y el ciclo celular, exponga una interpretación del gráfico adjunto.
Se han representado los diferentes estados del cromosoma a lo largo del ciclo celular. Se puede interpretar que los dibujos A y B corresponden a la interfase y los restantes a la fase M.
El dibujo C corresponde a la metafase, D y E a la anafase y F, a la telofase.
El dibujo A corresponde al período G1 de la interfase. En el B se observa que ha empezado la replicación o síntesis de ADN (fase S).
11.- Interprete esta otra versión del estado del cromosoma durante el ciclo celular. ¿Qué conclusión puede aportar?
Se puede interpretar que los dibujos A y B corresponden a la fase M y los otros tres, a la interfase.
Fase M (división celular): A = metafase. B = anafase.
Interfase: C = G1. D = fase S. E = G2.
Conclusión: el cromosoma está formado por una sola molécula de ADN, desde la anafase hasta el comienzo de la fase S, o por dos, desde el final de S hasta el inicio de la anafase.
Nota.- El dibujo E también se podría considerar como profase.
12.- ¿Cómo es el ciclo celular en las células cancerosas? En relación con la quimioterapia, ¿a qué se llama cronoterapia?
En las células cancerosas está alterado el control normal de la división. Estas células crecen y se dividen de modo incesante, invadiendo otros tejidos, pues no presentan el fenómeno de la inhibición por contacto (frenar la división al contactar con otras células). En los cultivos de células normales, pasado un cierto tiempo, tras realizar un número determinado de divisiones, toda la población celular deja de dividirse y el cultivo envejece y muere. Por el contrario, el cultivo de células cancerosas crece indefinidamente.
Ensayando con dos tipos de cáncer hematológico se ha descubierto que las células tumorales carecen de una molécula encargada de regular el llamado ritmo circadiano, esto es, nuestro reloj biológico interno que regula los ciclos de sueño y vigilia. La molécula del reloj biológico que está inactiva en el cáncer es BMAL1, encargada de controlar que ciertos genes se expresen de forma rítmica. Por esta razón las células cancerosas estarían permanentemente despiertas (activas).
Este descubrimiento podría tener implicaciones a la hora de diseñar estrategias de tratamiento para los pacientes, administrándoles la quimioterapia en el momento del día más adecuado para que las células malignas respondan mejor (cronoterapia).
Así, pues, la cronoterapia consiste en hallar el momento para optimizar la aplicación del tratamiento con quimioterapia.
13.- ¿A qué se llama “senescencia replicativa”?
Las células de mamífero no se dividen infinitamente sino un número limitado de veces antes de diferenciarse en células altamente especializadas. Por ejemplo, los fibroblastos humanos en un medio de cultivo estándar se dividen entre 25 y 50 veces.
Se ha observado que la proliferación celular va disminuyendo su velocidad y finalmente se detiene, fenómeno que se ha denominado “senescencia replicativa”.
14.- ¿Qué característica del ciclo celular presenta una mayor relevancia?
Una de las características esenciales del ciclo celular es la existencia de mecanismos de control para conocer la fiabilidad del proceso y bloquearlo en el caso de que no se hayan realizado adecuadamente todos los procesos. Estos puntos de control garantizan, principalmente, la replicación y reparación del ADN así como la entrada y salida de la mitosis.
15.- ¿Cuántos puntos de control suelen considerarse en el ciclo celular? ¿Qué aspectos son supervisados en las fases G1, G2 y M?
Se han postulado cuatro puntos en los que se controla el estado de la célula y del medio extracelular, con objeto de decidir si se realizan o restringen las acciones propias de cada una de las fases del ciclo. Estos cuatro puntos de control son: el de restricción (R) y, sobrepasado éste, otros tres ubicados al final de G1, de G2 y en la fase M.
El primer punto de control se encuentra en G1, justo después del punto de restricción. Aquí se controla el tamaño celular y si las condiciones del entorno son favorables.
Al final de G2 (2º punto de control) se controla si está replicado todo el ADN, si es favorable el entorno y si el tamaño celular es apropiado para la división.
El 3er punto de control se halla en la fase M, entre la metafase y anafase. Se controla principalmente si todos los cromosomas están unidos al huso mitótico, así como si están alineados en el ecuador celular. Si se detecta que uno de los cinetocoros no se encuentra unido a los microtúbulos, se genera una señal al sistema de control que bloquea la activación de proteínas implicadas en la separación de las cromátidas hermanas.
16.- ¿Es de carácter extracelular el control del ciclo celular?
El control del ciclo celular se presenta a dos niveles, intracelular y extracelular.
Las células en los organismos pluricelulares proliferan solo cuando se requieren más células. La entrada al ciclo celular no es un proceso autónomo de la célula, pues se requiere la presencia de señales adecuadas, esto es, factores solubles de naturaleza proteica (mitógenos), procedentes del medio extracelular o de otras células.
17.- ¿Cómo se ejerce a nivel intracelular el control del ciclo?
El control del ciclo a nivel intracelular se realiza previa fosforilación, mediante quinasas específicas, de unas proteínas clave para posibilitar la replicación, la mitosis y la citocinesis.
La actividad de estas quinasas, llamadas Cdk (Cyclin dependent kinases) depende de unas proteínas activadoras llamadas ciclinas, que se sintetizan en determinados momentos del ciclo celular y posteriormente se degradan (el número de ciclinas identificadas es distinto en eucariotas inferiores y superiores).
En resumen: la regulación a nivel intracelular es llevada a cabo por complejos Cdk-ciclinas y sus inhibidores, entre otras proteínas. El fundamento del control del ciclo celular radica esencialmente en los distintos complejos Cdk-ciclinas, que controlan el inicio de las fases S y M.
19.- ¿Qué relación tiene la proteína p27 con el ciclo celular?
La proteína p27 (masa molecular 27 kDa, esto es, 27000 u) se une a ciclinas y Cdk inactivando el complejo ciclina-cdk, circunstancia que bloquea la entrada en la fase S.
20.- ¿Qué relación existe entre la proteína p53 y el ciclo celular?
La proteína p53 (masa molecular 53000 u, o sea, 53 kDa) es un regulador de la proliferación de la mayoría de los tipos celulares. Se trata de un factor de transcripción que, al fijarse en una secuencia concreta del ADN, activa los genes implicados en el programa de apoptosis o en la detención del ciclo celular. Concretamente, la inhibición de la proteína p53 provoca una proliferación celular.
21.- ¿Cómo actúa la p53?
Cuando el ADN presenta un daño limitado, aumenta la concentración de proteína p53. Este aumento desencadena una serie de reacciones y estimula la síntesis de otra proteína, llamada p21, la cual, por su acción inhibidora, provoca la inactivación de un complejo ciclina-Cdk (Cdk2-Ciclina E) y la consiguiente detención del ciclo.
Esta situación proporciona tiempo para la actividad de los sistemas de reparación del ADN, pues mientras persista la inactivación de dicho complejo la célula no puede entrar en la fase S.
Cuando el ADN es reparado, la proteína p53 se libera de la región promotora del gen p21, provocando el descenso en los niveles de p21. Esto permite restaurar la actividad del complejo ciclina-Cdk y la continuación del ciclo.
Nota.- La p53 es una proteína integrada por 393 aminoácidos y, estructuralmente, presenta 3 dominios. Es inestable en condiciones normales porque se encuentra unida a otra proteína, que funciona como un “marcador”, para que la citada p53 se degrade. El incremento de la concentración de p53 requiere la previa separación de su “marcador”, llevada a cabo por otros mediadores.
En células normales, el nivel de la proteína p53 es bajo porque es ubiquitinada y destruida por Mdm2. Los daños del ADN y otras señales de estrés pueden hacer que no se una a Mdm2 e incrementar su concentración, activando la transcripción del gen p21.
22.- ¿Qué significado tiene la expresión “el guardián del genoma”?
Cuando el ADN sufre daño, el gen p53, que codifica a la proteína p53, se activa y el incremento de la misma desencadena la ralentización o detención del ciclo celular, dando tiempo para que actúe el sistema de reparación de ADN, razón por la cual se ha denominado “el guardián del genoma”.
El gen p53 se halla en el brazo corto del cromosoma 17 y activa su expresión en situación de estrés o daño celular. Bloquea la proliferación o división de la célula alterada e induce la apoptosis (suicidio celular).
Nota.- Cuando fue descubierto, en 1979, el gen p53 se consideró dentro de los oncogenes (genes que promueven el cáncer). Pero, en 1989, pasó a integrar el grupo de los supresores tumorales. Ello se debió a que su primera identificación se obtuvo de una de sus formas mutadas y no de la normal o natural.
23.- ¿Qué son los genes supresores de tumores?
Ciertas proteínas, llamadas inhibidores de Cdk, y algunos factores de transcripción, como la proteína p53, tienen la función de impedir la proliferación celular. Los genes que codifican estas proteínas se llamaron “genes supresores de tumores”, como por ejemplo, el gen p53.
La mutación de los genes que codifican dichas proteínas, o bien, la disfunción de las mismas, implica la pérdida de control del ciclo celular y la incapacidad para detenerlo, lo cual conduce a la proliferación celular (tumorogénesis).
Las células que presentan los dos alelos del gen p53 mutados, no tendrán proteína p53 funcional y, por consiguiente, continuarán dividiéndose a pesar del daño en su genoma, circunstancia que, en general, desencadenará un proceso canceroso.
24.- Enumere las bases conceptuales del ciclo celular.
Fueron las contribuciones de los científicos galardonados con el Premio Nobel de Medicina de 2001 las que sirvieron para establecer las bases conceptuales del ciclo celular, que podemos exponer en cinco puntos:
1) El ciclo celular puede considerarse como una secuencia de acontecimientos organizados temporalmente.
2) El inicio de cada uno de ellos es consecuencia de la finalización de los anteriores.
3) La relación entre unos acontecimientos y otros se establece directamente o a través de algún tipo de señales.
4) La finalización de una etapa antes de comenzar la siguiente conlleva la existencia de controles a lo largo del ciclo.
5) Ciertos acontecimientos actúan como limitantes para la progresión del ciclo
25.- Resuma las características del ciclo celular.
• El ciclo celular es un conjunto de procesos ordenados que lleva a cabo la célula que va a dividirse. Comprende la interfase y la división celular.
• El control del ciclo celular se presenta a dos niveles, intracelular y extracelular.
• El control intracelular está a cargo de mediadores proteicos que ejercen una acción activadora o inhibidora.
• Hay un punto de restricción y tres puntos de control, regulados por distintas combinaciones de Cdk-ciclinas.
• La entrada al ciclo celular requiere señales adecuadas (mitógenos), procedentes del medio extracelular o de otras células.
• Cuando una célula no es necesaria o se convierte en una posible amenaza por estar infectada o alterada, puede morir por necrosis o apoptosis.
2.- Defina el concepto de apoptosis.
Apoptosis se puede definir como la muerte fisiológica de las células.
Aunque dicho término se considera sinónimo de muerte celular programada, la desaparición de muchas células no está prefijada, sino que es consecuencia de cambios impredecibles en su entorno. La apoptosis es generalmente inducida por agentes externos a la célula, con frecuencia producidos por otras células del organismo. Son las señales de pervivencia, de naturaleza cambiante en distintos períodos, las que impiden el inicio del suicidio celular. La regla general es que el destino de la célula no sólo depende del tipo de señales que reciba en cada instante, sino también de las que haya recibido antes (una excepción a esta regla es la actuación de los linfocitos citotóxicos).
La expresión “muerte celular programada” es en cierto modo equívoca, pues sugiere que la célula tiene fijada genéticamente una fecha de muerte. Lo correcto es interpretar que la célula posee un programa de autodestrucción, pero que sólo será ejecutado ante señales o situaciones muy concretas. Por esta razón se ha propuesto la denominación de suicidio inducido.
Nota.- El término apoptosis deriva del griego arcaico y alude a la caída natural de las hojas. Fue acuñado en 1972 por John Kerr y colaboradores.
3.- ¿Qué se requiere para desencadenar la apoptosis? ¿En qué momento del ciclo celular se toma la decisión de iniciar la apoptosis?
Las células de un ser pluricelular deben estar coordinadas mediante señales químicas para optimizar su fisiología. La respuesta a estas señales es selectiva e implica la recepción de la molécula señal mediante una proteína receptora específica, seguida de la correspondiente transducción, que transforma la señal extracelular en señales intracelulares que modificarán su actividad.
En ausencia de señales concretas o en presencia de señales “de muerte” la célula pone en marcha un programa que provoca su propia destrucción, teniendo lugar una degradación progresiva de los materiales y estructuras celulares, proceso denominado apoptosis o muerte celular controlada.
Existen proteínas extracelulares que, como las neurotrofinas, pueden favorecer la supervivencia, o por el contrario inducir la apoptosis, como ocurre con el ligando de Fas.
La decisión de proseguir el ciclo o entrar en la vía terminal de apoptosis se toma en el punto de control situado al final de la fase G1del ciclo celular.
4.- Haga un resumen sobre las diversas señales que regulan el suicidio celular.
La célula recibe señales de multiplicación celular que promueven la expresión de los llamados protooncogenes, que además de iniciar tal multiplicación, intervienen en la activación de las ciclinas para garantizar el curso de todas las etapas del ciclo celular.
Señales provenientes del medio extracelular, tales como el factor de necrosis tumoral (TNF) y el ligando del receptor Fas, inducen el suicidio celular.
La iniciación del suicidio se realiza mediante activación de las proteasas ICE.
Cuando la célula recibe menos señales de supervivencia disminuye la expresión de los genes inhibidores del suicidio, como Bcl-2.
5.- ¿Cómo actúan los linfocitos citotóxicos (o verdugos)?
Los linfocitos citotóxicos o verdugos, al entrar en contacto con las células infectadas, llevan a cabo la secreción de unos gránulos citoplásmicos que contienen proteínas citotóxicas: perforina y granzimas (las dos más importantes son la granzima A y la granzima B).
La perforina interacciona con las membranas celulares y permite el acceso de las granzimas al interior de la célula diana, las cuales inducen muerte celular por apoptosis.
Las granzimas desempeñan un papel crucial, pues intervienen en la activación proteolítica de las enzimas ICE de la célula alterada o infectada, desencadenando el suicidio de la misma, sin que en este caso influyan las señales que la célula reciba de su entorno.
Nota.- Algunos autores concluyen que el sistema perforina-granzima B induce apoptosis a través de la activación de caspasas, implicando a la vía mitocondrial, y que el sistema perforina-granzima A induce apoptosis a través de la generación de radicales libres, también por la vía mitocondrial, pero en ausencia de activación de caspasas
6.- ¿A qué se refiere el llamado “carácter altruista” de la apoptosis?
Los seres pluricelulares recurren a la apoptosis en muchos procesos fisiológicos y patológicos. El carácter altruista de la apoptosis alude a que la célula se autodestruye en beneficio del organismo. De esta forma, la apoptosis, también llamada muerte celular controlada, contribuye al desarrollo y bienestar del organismo.
Este programa de muerte celular ocurre, por ejemplo, en células infectadas por virus (evitando con ello la producción de nuevas partículas víricas) o en las que tengan importantes alteraciones en su ADN.
7.- ¿Qué significado tiene la frase: “La muerte celular forma parte de la vida”?
En la ontogénesis de cada ser vivo pluricelular, la muerte de ciertas células es necesaria para el correcto desarrollo orgánico.Concretamente, un embrión humano de pocas semanas tiene los dedos unidos por una membrana cuyas células se eliminan a sí mismas, dejando los dedos bien separados.
Además, todo ser vivo debe desprenderse de las células que a lo largo de su vida han ido acumulando mutaciones y errores susceptibles de convertirlas en cancerosas. Cualquier desarreglo que afecte a la capacidad de morir de estas células puede tener consecuencias letales para el organismo.
8.- ¿A qué se refiere la frase: “La muerte es escultora de vida”?
A lo largo del desarrollo embrionario la muerte celular modela la forma del embrión eliminando tejidos, como si fuese un escultor. Así, por ejemplo, la cola y las branquias del renacuajo desaparecen durante la metamorfosis en rana.
En el caso humano, la cola del embrión también es eliminada gracias a la muerte celular, quedando esculpido un feto sin cola, y la formación de la mano, mediante eliminación del tejido interdigital, ya se ha mencionado en la cuestión anterior.
9.- ¿Es lo mismo necrosis celular que apoptosis? ¿Qué factores pueden causar necrosis?
La necrosis o muerte accidental se produce cuando la célula sufre daño grave. La célula lesionada se hincha y la membrana celular se rompe, expulsando al medio su contenido, situación que suele desencadenar un proceso inflamatorio y provocar un daño más o menos severo en la matriz intercelular y en las células vecinas.
Por el contrario, en la apoptosis no hay expulsión de materiales celulares y, por lo general, las células que mueren de esta forma son células sanas, esto es, potencialmente funcionales, a diferencia de las que mueren por necrosis.
En la necrosis las células pueden ser alteradas por una serie de factores: lesiones, hipoxia, toxinas, agentes infecciosos, etc.
10.- ¿En qué consiste la apoptosis o muerte celular programada (MCP) y cuáles son las características morfológicas?
Actualmente se consideran tres formas o modalidades de muerte celular programada (MCP): tipo 1, apoptosis, que es la más común; tipo 2, autofagia, mediante acción de los lisosomas; tipo 3, muerte vesicular no lisosomal.
La apoptosis es el tipo de MCP más frecuente y consiste, fundamentalmente, en la activación de las enzimas ejecutoras del programa. Se trata de una muerte “natural” y las células se autodestruyen en un período de tiempo que en general oscila alrededor de 60 minutos, pero sin expulsar al medio su contenido.
Morfológicamente se caracteriza por: retracción celular, disminución de la turgencia, desorganización del citoesqueleto y de la lámina nuclear, invaginación de la membrana nuclear, fragmentación del material cromosómico (la rotura del ADN en grandes fragmentos sería la señal irreversible que conduce a la apoptosis), formación de protuberancias en la superficie celular. Finalmente, la célula origina varias vesículas o cuerpos apoptóticos (con orgánulos y fragmentos nucleares), que en general son fagocitados por los macrófagos.
11.- ¿Qué relación hay entre la apoptosis y las mitocondrias?
Las mitocondrias son los orgánulos que producen la energía de la célula, por lo que cualquier disfunción conduce a la desregulación del medio intracelular. En tales circunstancias la mitocondria deja de producir eficientemente energía y libera una serie de moléculas que pueden desencadenar la apoptosis.
Una de estas moléculas es el citocromo c, proteína crucial en la transferencia de electrones propia de la respiración celular, pero que en el citosol activa la vía de las caspasas.
La mitocondria puede liberar también el factor inductor de la apoptosis (AIF), una proteína cuya actividad apoptótica pasa por su transporte al núcleo y es independiente, al menos en parte, de la vía de las caspasas.
15.- ¿Por qué se dice que el gen p53 puede desempeñar el papel de policía o de asesino?
Este gen vela por la integridad del genoma y obliga a la célula a interrumpir su multiplicación y a reparar su lesión genética cuando ésta se produce, es decir, detiene el ciclo celular, actuando como policía, hasta que se termina la reparación, razón por la cual fue denominado “guardián” del genoma.
Las mutaciones que provocan la disfunción del gen p53 tienen como consecuencia que las células alteradas puedan seguir multiplicándose y sufrir nuevas transformaciones genéticas favorables a la tumoración (en torno al 50 % de los casos de cáncer en humanos presentan mutaciones en dicho gen).
Por otra parte, el gen p53 participa en la apoptosis o muerte celular programada, conduciendo a las células al suicidio cuando el daño en el ADN es tan significativo que rebasa la capacidad de reparación. Así, pues, cuando la lesión en el ADN es demasiado importante, el gen p53 desempeña el papel de asesino puesto que desencadena la muerte de la célula.
Nota.- El gen p53 está localizado en el brazo corto del cromosoma 17 y codifica a la proteína p53, cuya concentración intracelular debe estar bien regulada, pues si el nivel es demasiado alto puede exacerbar la apoptosis. El regulador principal de p53 es Mdm2, que actúa “marcando” a p53 para que sea ubiquitinada y degradada.
La proteína p53 es capaz de unirse a lugares específicos del ADN, regulando la expresión de otros genes que controlan funciones celulares esenciales. Esta es la razón de la conexión del gen p53 con procesos tales como la carcinogénesis, el ciclo celular y la activación de la apoptosis.
16.- ¿Cuál es la probable relación entre una mutación temprana del gen p53 y el desarrollo del cáncer de pulmón?
La proteína p53 (codificada por el gen p53), que bloquea el ciclo celular cuando el ADN está alterado, puede llegar a inducir apoptosis en la célula cuyo ADN esté severamente dañado. La mutación temprana del gen p53 tenderá a impedir que células anormales mueran por apoptosis, dejando vía libre al proceso de tumorogénesis.
17.- Busque en Internet el artículo “Apoptosis, señalización celular y transformaciones tumorales en Drosophila” (Ginés Morata. Centro de Biología Molecular “Severo Ochoa”, CSIC-UAM, Madrid). Encuentre la respuesta a las siguientes cuestiones:
A) ¿Qué funciones cumple la apoptosis?
B) ¿Cómo se puede inducir apoptosis?
C) ¿Es posible mantener vivas durante mucho tiempo células apoptóticas?
A) La mortalidad celular programada, o apoptosis, cumple varias funciones muy importantes durante el desarrollo animal.
Es necesaria para muchos procesos morfogenéticos, así como para impedir que haya exceso de células o eliminar células dañadas o debilitadas durante el desarrollo.
La apoptosis ha de estar regulada de forma muy precisa porque su desregulación da lugar frecuentemente a procesos tumorales y su exceso está relacionado con la aparición de enfermedades neurodegenerativas en humanos.
B) Existen órganos, como el disco imaginal de ala de Drosophila, donde no hay apoptosis en el desarrollo normal. Sin embargo, este proceso se puede inducir mediante tratamientos de estrés con irradiación (rayos X) o choque térmico, siempre que se haya inhibido previamente la actividad caspasa.
En estas condiciones se ha observado que las células emiten las moléculas señalizadoras Decapentaplegic (Dpp) y Wingless (Wg), las cuales inducen proliferación adicional en las células vecinas. Dpp y Wg son los morfógenos principales en el desarrollo deDrosophila, y sus homólogos en vertebrados (TFG-ß/BMP y Wnt) tienen funciones similares.
En experimentos recientes se ha estudiado el papel específico de las señales Dpp y Wg en el proceso, descubriendo que el estímulo proliferativo se debe únicamente a Dpp mientras que Wg tiene una actividad inhibitoria destinada a regular el efecto de Dpp.
C) También se ha descubierto, en experimentos con Drosophila, que se pueden mantener vivas indefinidamente las células apoptóticas mediante el uso de un inhibidor general de la actividad caspasa, la proteína de baculovirus P35.
19.- ¿Qué genes regulan la muerte celular de los organismos más complejos que Caenorhabditis elegans?
Relacionados con la regulación de la muerte celular, los seres humanos poseen dos genes muy parecidos a los del nematodo, los llamados ICE y Bcl-2, cuyas familias se componen al menos de 6 miembros cada una.
La expresión de los genes de la familia ICE promueve la apoptosis.
En la otra familia hay genes antagonistas, pues Bcl-2 y Bcl-XL favorecen la supervivencia impidiendo la activación o los efectos de las proteasas ICE, mientras que, por el contrario, Bax, Bad, Bag y Bak promueven el suicidio, oponiéndose a los efectos de los otros dos de su propia familia y apoyando la actividad de las citadas proteasas.
20.- ¿Está de acuerdo con la existencia de una cierta especialización de los genes reguladores de la muerte celular?
Se ha descubierto que la supervivencia de una población celular concreta viene asegurada en momentos diferentes de su existencia por la expresión de distintos genes de la familia Bcl-2. La expresión de Bcl-XL es indispensable, por ejemplo, para la supervivencia de los linfocitos embrionarios, mientras que la de Bcl-2 lo es para la de los linfocitos adultos.
A esta especialización de los genes reguladores de la muerte celular se superpone la diversidad de las reacciones que presentan las células ante determinada señal. Puesto que una misma señal puede desencadenar la expresión de diferentes genes en poblaciones celulares distintas, provocará la muerte de unas y la supervivencia de otras, circunstancia que apunta hacia la enorme complejidad de la regulación genética y molecular del suicidio celular.
21.- ¿Cómo se comportan las células normales y las cancerosas cuando emigran a otros órganos?
Una célula normal que emigre a un órgano que no es el suyo tendrá serios problemas de supervivencia, puesto que allí no recibe las señales necesarias para impedir que actúe su programa de autodestrucción.
Por el contrario, una célula cancerosa, cuyo programa de suicidio está inutilizado, puede seguir viviendo en ausencia de las señales apropiadas, circunstancia que permite colonizar nuevos órganos y formar metástasis.
22.- ¿Hay apoptosis en los procesos de tumorogénesis?
Se dan todos los grados intermedios entre los tumores cuyas células mueren por apoptosis en número considerable, pero se multiplican aún más aprisa, y aquellos cuyas células se multiplican prácticamente sin freno apoptótico, que son los más nocivos y los que en el caso humano suelen tener un pronóstico infausto.
OTROS ENLACES CON PREGUNTAS RELACIONADAS:
No hay comentarios:
Publicar un comentario